

GPCRs in Cancer: Protease-Activated Receptors, Endocytic Adaptors and Signaling
- PMID: 29954076
- PMCID: PMC6073120
- DOI: 10.3390/ijms19071886
GPCRs in Cancer: Protease-Activated Receptors, Endocytic Adaptors and Signaling
Abstract
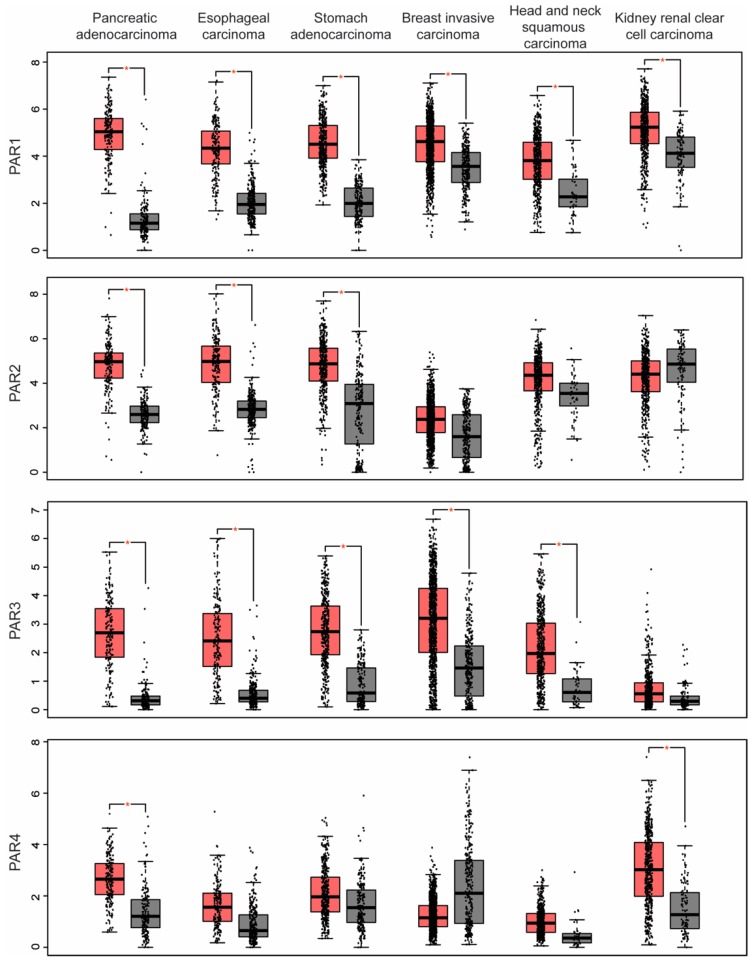
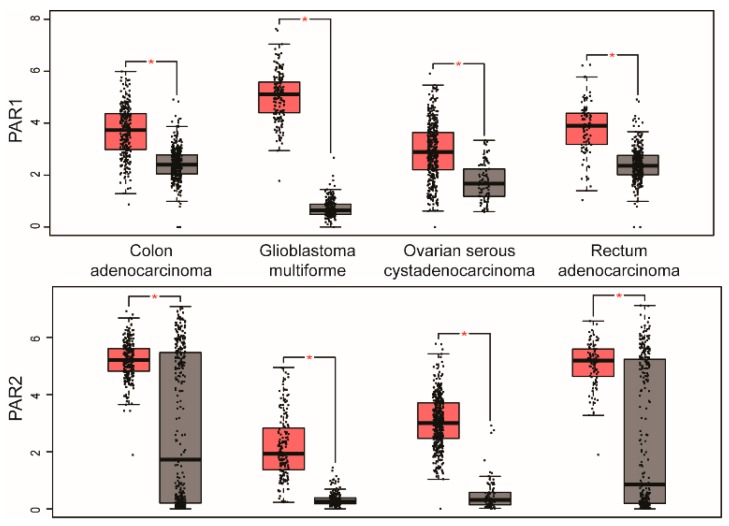
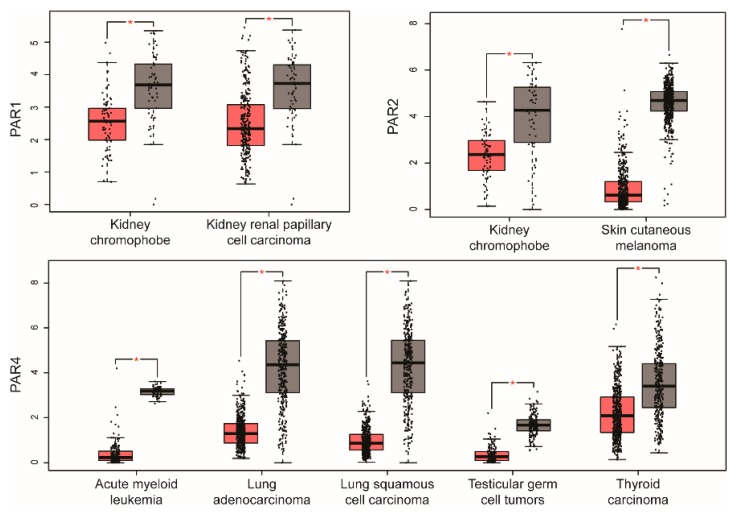
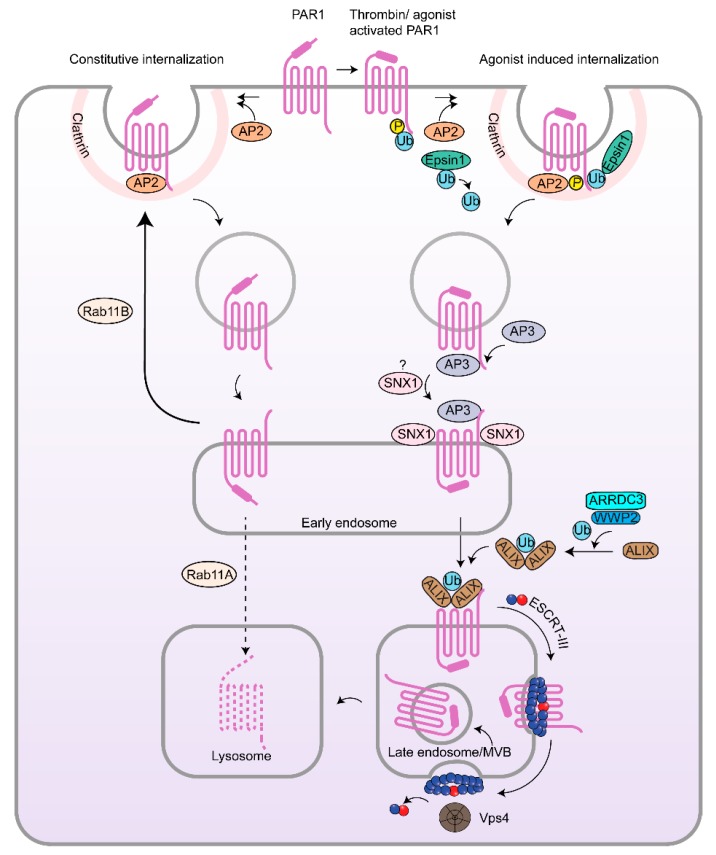
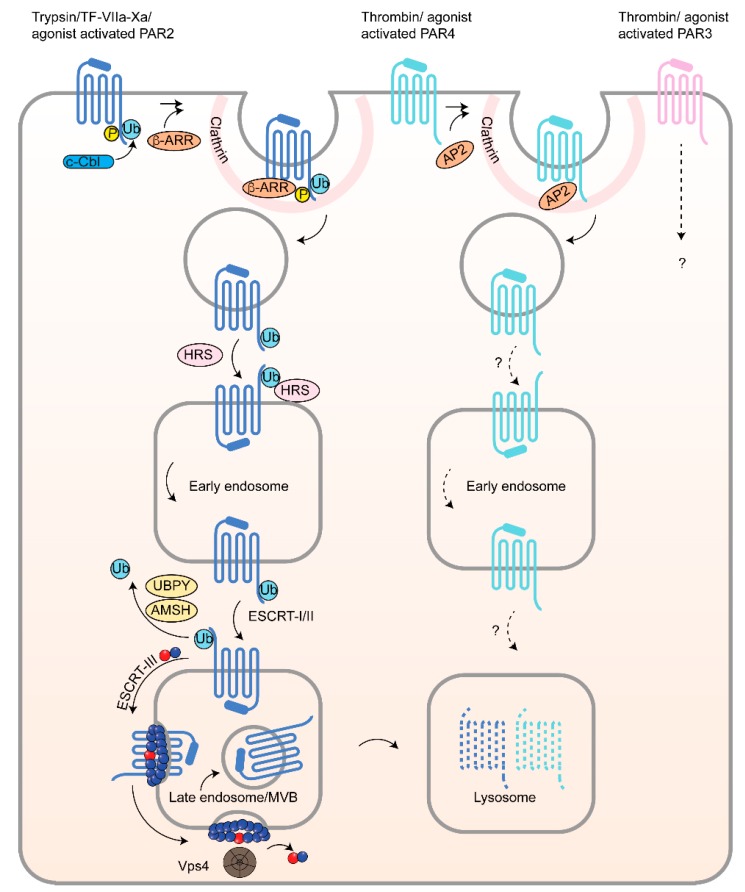
G protein-coupled receptors (GPCRs) are a large diverse family of cell surface signaling receptors implicated in various types of cancers. Several studies indicate that GPCRs control many aspects of cancer progression including tumor growth, invasion, migration, survival and metastasis. While it is known that GPCR activity can be altered in cancer through aberrant overexpression, gain-of-function activating mutations, and increased production and secretion of agonists, the precise mechanisms of how GPCRs contribute to cancer progression remains elusive. Protease-activated receptors (PARs) are a unique class of GPCRs implicated in cancer. PARs are a subfamily of GPCRs comprised of four members that are irreversibly activated by proteolytic cleavage induced by various proteases generated in the tumor microenvironment. Given the unusual proteolytic irreversible activation of PARs, expression of receptors at the cell surface is a key feature that influences signaling responses and is exquisitely controlled by endocytic adaptor proteins. Here, we discuss new survey data from the Cancer Genome Atlas and the Genotype-Tissue Expression projects analysis of expression of all PAR family member expression in human tumor samples as well as the role and function of the endocytic sorting machinery that controls PAR expression and signaling of PARs in normal cells and in cancer.
Keywords: ARRDC3; arrestins; breast cancer; invasion; lysosomes; metastasis.
Conflict of interest statement
This manuscript reports unpublished work that is not under consideration for publication elsewhere. All co-authors on this work agree to its content and hereby declare no competing commercial interests relating to this submitted work.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
