

Pore properties of Orai1 calcium channel dimers and their activation by the STIM1 ER calcium sensor
- PMID: 29954946
- PMCID: PMC6102135
- DOI: 10.1074/jbc.RA118.003424
Pore properties of Orai1 calcium channel dimers and their activation by the STIM1 ER calcium sensor
Abstract
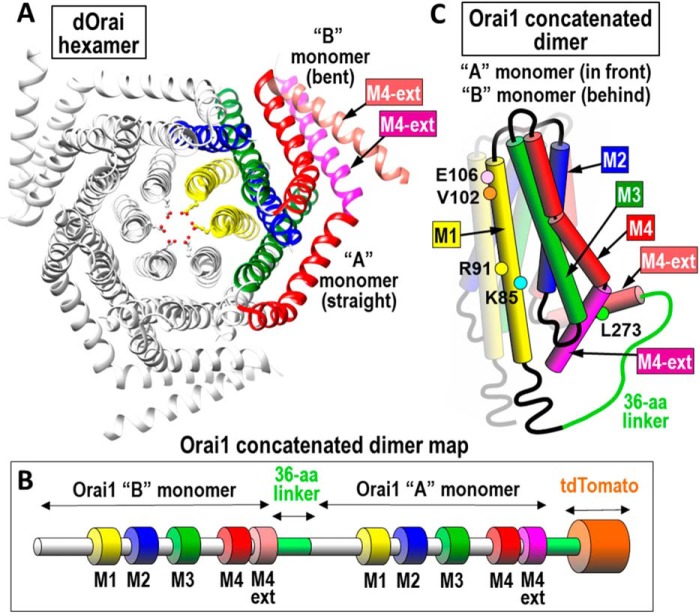
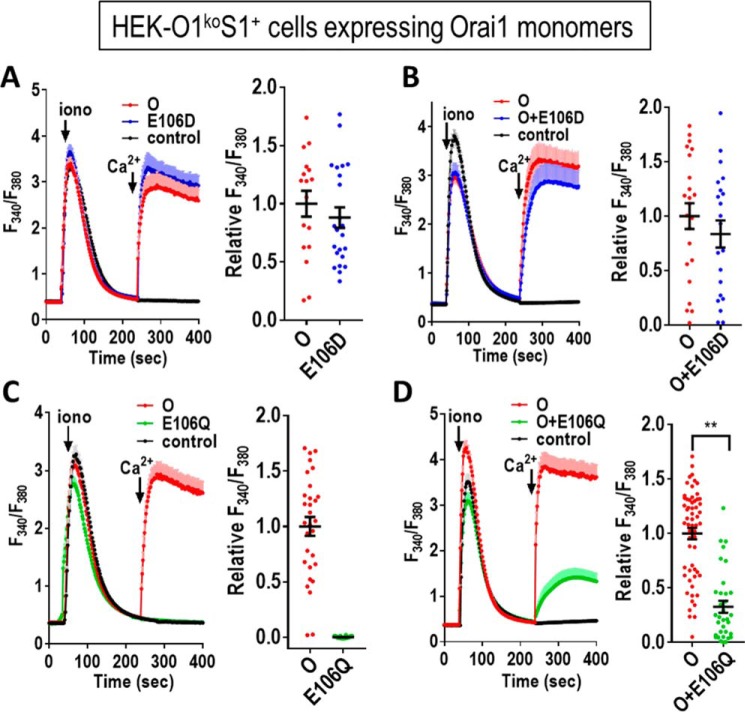
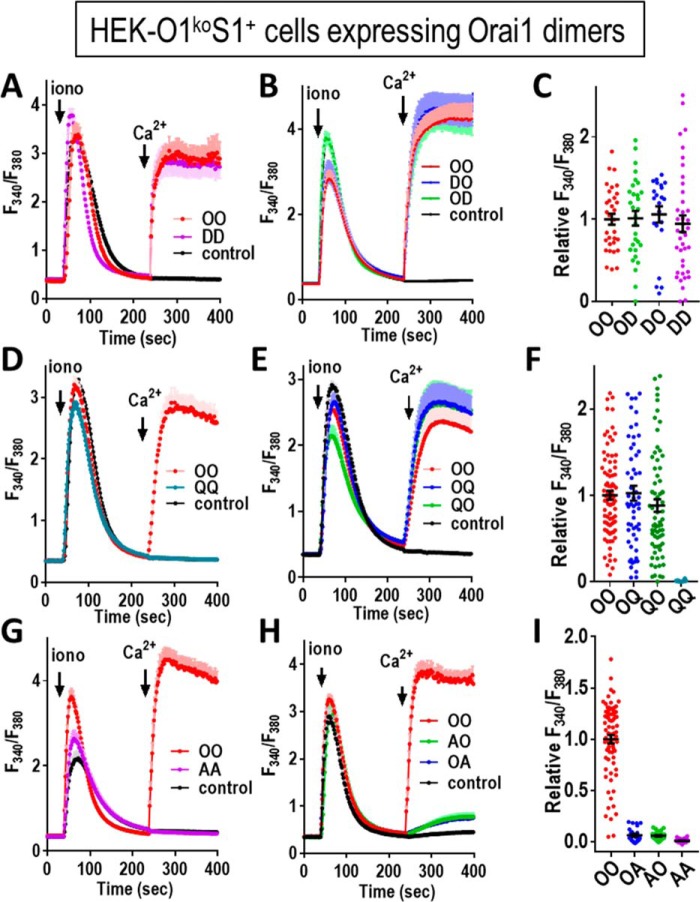
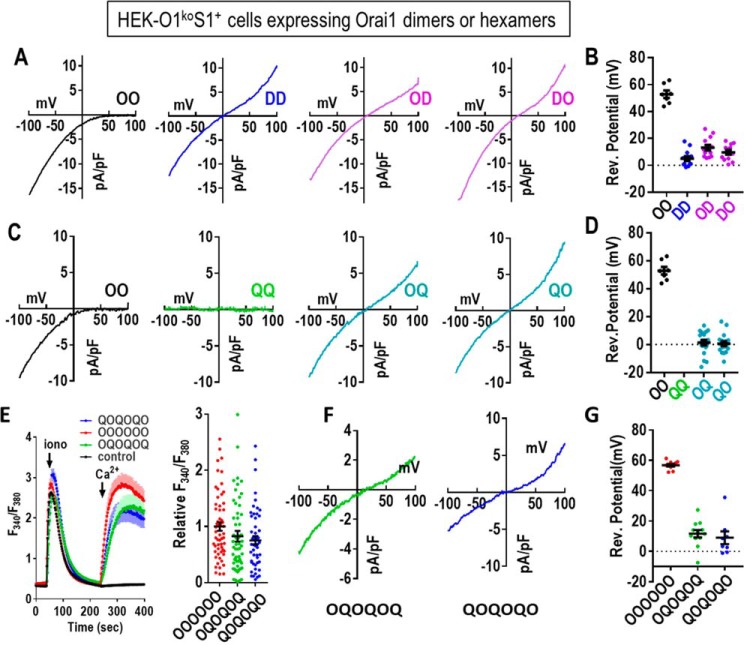
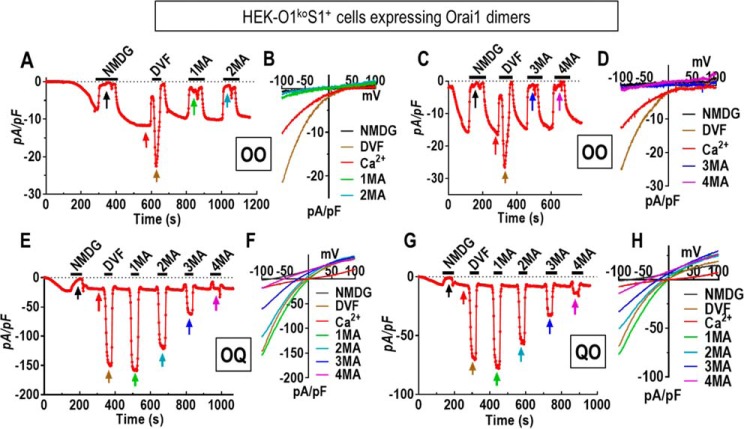
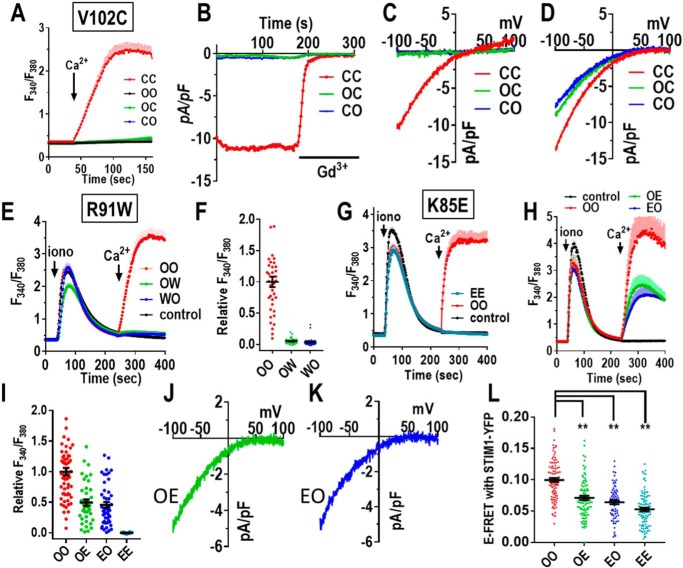
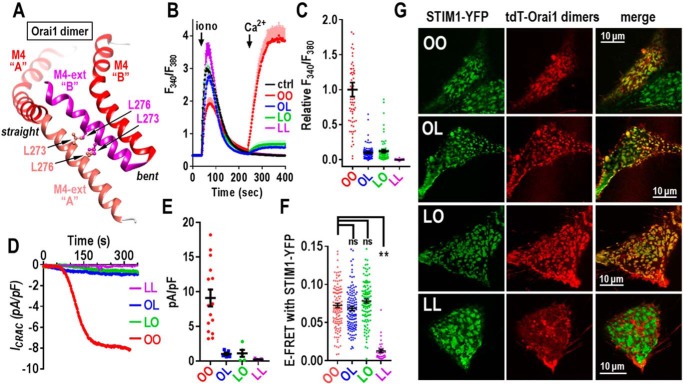
Store-operated Ca2+ entry signals are mediated by plasma membrane Orai channels activated through intermembrane coupling with Ca2+-sensing STIM proteins in the endoplasmic reticulum (ER). The nature of this elaborate Orai-gating mechanism has remained enigmatic. Based on the Drosophila Orai structure, mammalian Orai1 channels are hexamers comprising three dimeric subunit pairs. We utilized concatenated Orai1 dimers to probe the function of key domains within the channel pore and gating regions. The Orai1-E106Q selectivity-filter mutant, widely considered a dominant pore blocker, was surprisingly nondominant within concatenated heterodimers with Orai1-WT. The Orai1-E106Q/WT heterodimer formed STIM1-activated nonselective cation channels with significantly enlarged apparent pore diameter. Other Glu-106 substitutions entirely blocked the function of heterodimers with Orai1-WT. The hydrophobic pore-lining mutation V102C, which constitutively opens channels, was suppressed by Orai1-WT in the heterodimer. In contrast, the naturally occurring R91W pore-lining mutation associated with human immunodeficiency was completely dominant-negative over Orai-WT in heterodimers. Heterodimers containing the inhibitory K85E mutation extending outward from the pore helix gave an interesting partial effect on both channel activation and STIM1 binding, indicating an important allosteric link between the cytosolic Orai1 domains. The Orai1 C-terminal STIM1-binding domain mutation L273D powerfully blocked STIM1-induced channel activation. The Orai1-L273D/WT heterodimer had drastically impaired STIM1-induced channel gating but, unexpectedly, retained full STIM1 binding. This reveals the critical role of Leu-273 in transducing the STIM1-binding signal into the allosteric conformational change that initiates channel gating. Overall, our results provide important new insights into the role of key functional domains that mediate STIM1-induced gating of the Orai1 channel.
Keywords: Orai1; STIM1; calcium; calcium channel; cell signaling; channel gating; ion channel; selectivity filter; store-operated channel.
© 2018 Cai et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
