

Go/No-Go task engagement enhances population representation of target stimuli in primary auditory cortex
- PMID: 29955046
- PMCID: PMC6023878
- DOI: 10.1038/s41467-018-04839-9
Go/No-Go task engagement enhances population representation of target stimuli in primary auditory cortex
Abstract
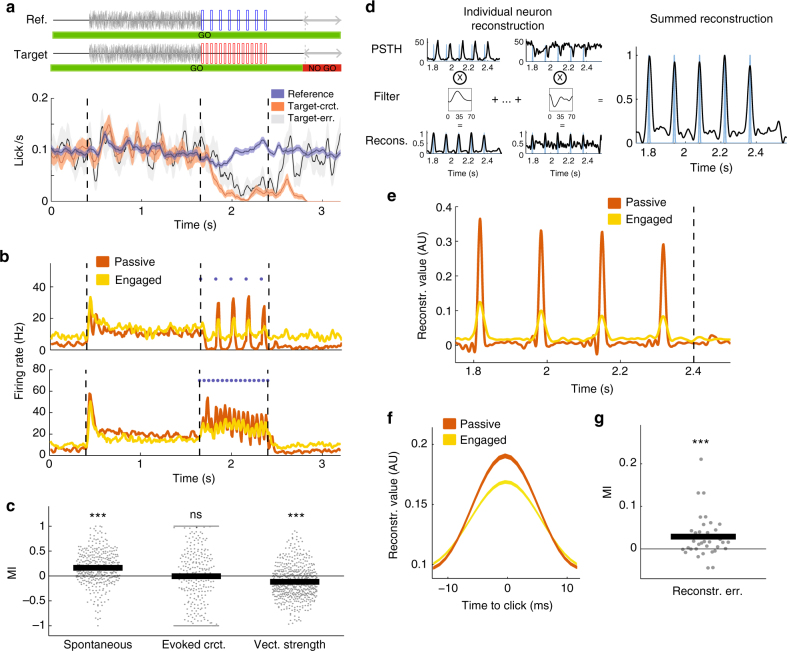
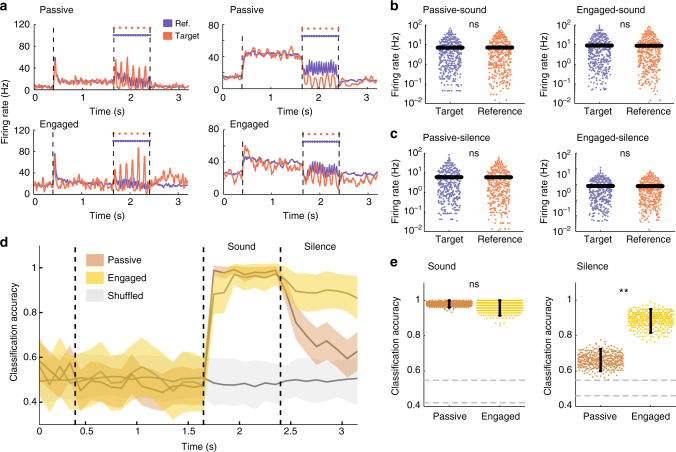
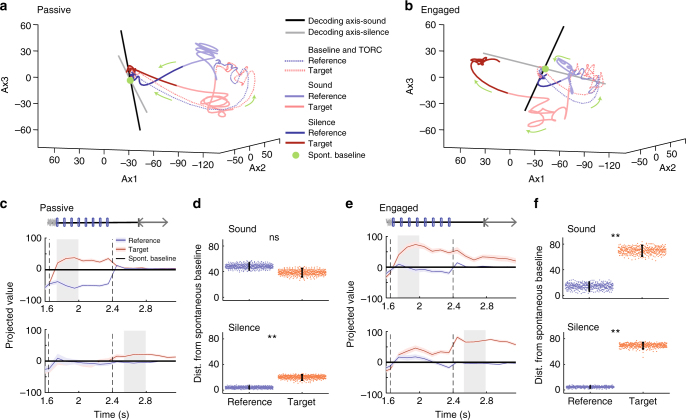
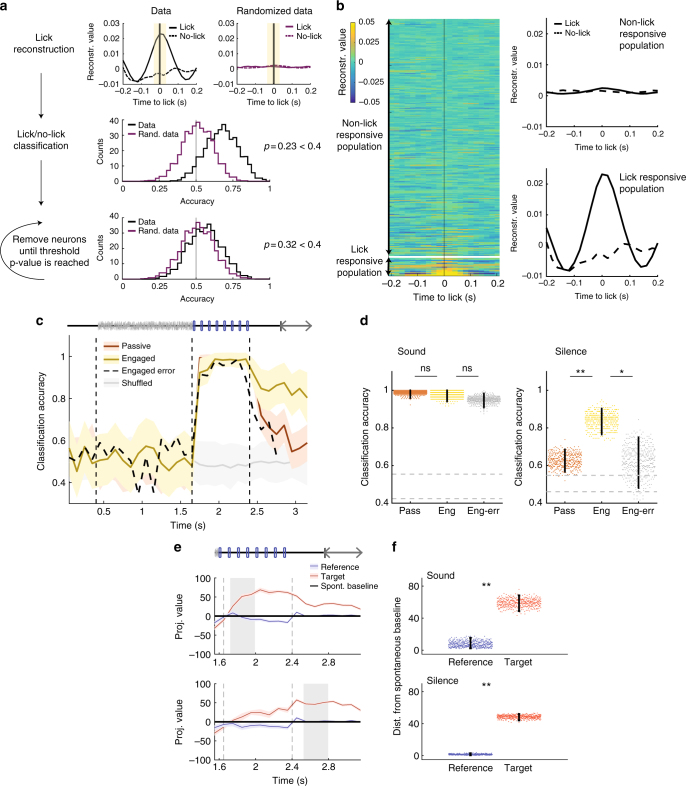
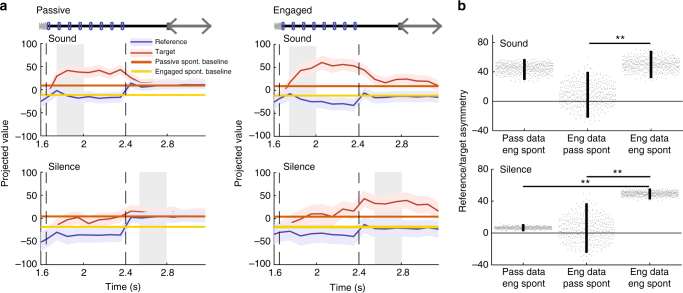
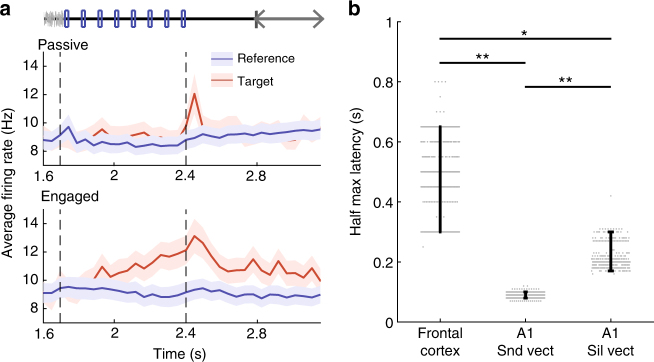
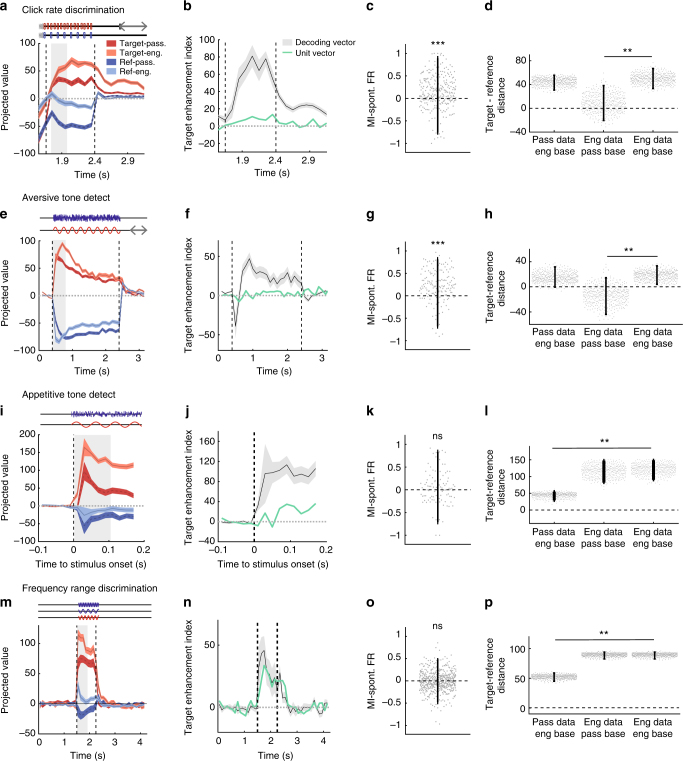
Primary sensory cortices are classically considered to extract and represent stimulus features, while association and higher-order areas are thought to carry information about stimulus meaning. Here we show that this information can in fact be found in the neuronal population code of the primary auditory cortex (A1). A1 activity was recorded in awake ferrets while they either passively listened or actively discriminated stimuli in a range of Go/No-Go paradigms, with different sounds and reinforcements. Population-level dimensionality reduction techniques reveal that task engagement induces a shift in stimulus encoding from a sensory to a behaviorally driven representation that specifically enhances the target stimulus in all paradigms. This shift partly relies on task-engagement-induced changes in spontaneous activity. Altogether, we show that A1 population activity bears strong similarities to frontal cortex responses. These findings indicate that primary sensory cortices implement a crucial change in the structure of population activity to extract task-relevant information during behavior.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Grants and funding
- R01 DC014950/DC/NIDCD NIH HHS/United States
- ANR-10-IDEX-0001-02 PSL*/EC | European Research Council (ERC)/International
- ANR-10-IDEX-0001-02 PSL*/Agence Nationale de la Recherche (French National Research Agency)/International
- ANR-10-LABX-0087 IEC/Agence Nationale de la Recherche (French National Research Agency)/International
LinkOut - more resources
Full Text Sources
Other Literature Sources
