

Duplication of hsp-110 Is Implicated in Differential Success of Globodera Species under Climate Change
- PMID: 29955862
- PMCID: PMC6188557
- DOI: 10.1093/molbev/msy132
Duplication of hsp-110 Is Implicated in Differential Success of Globodera Species under Climate Change
Abstract
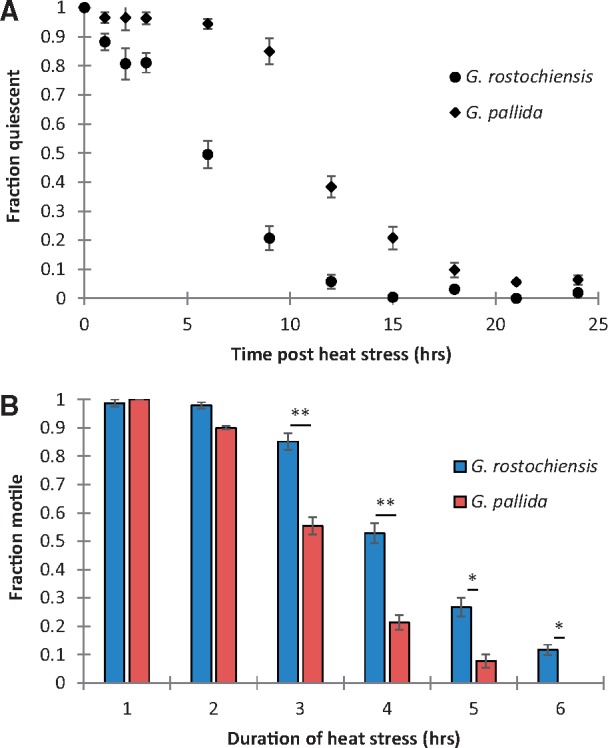
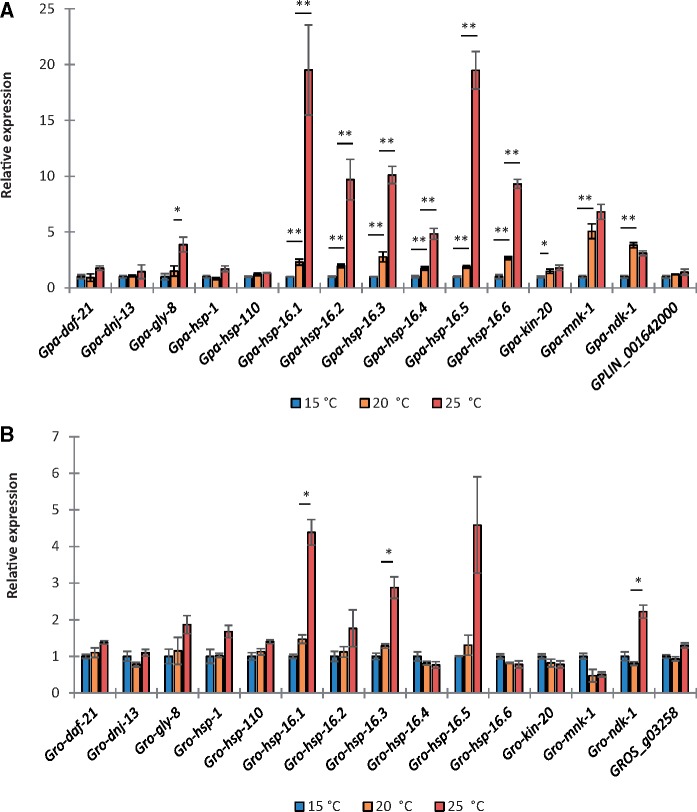
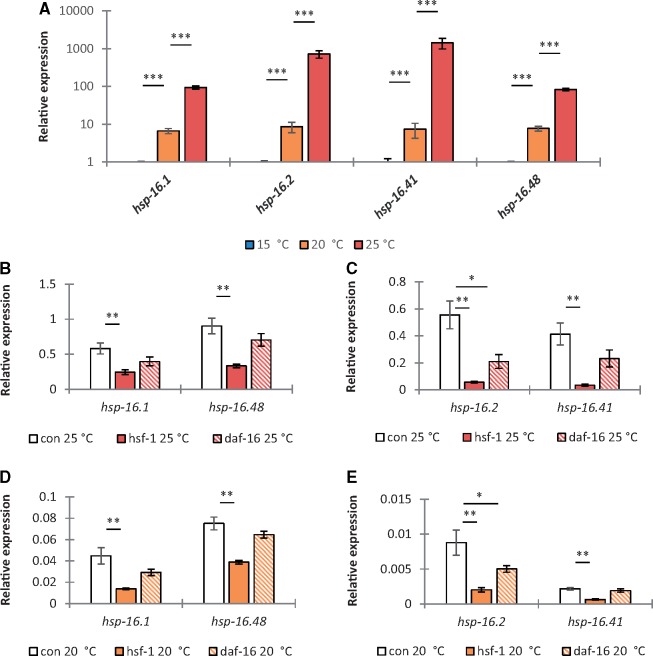
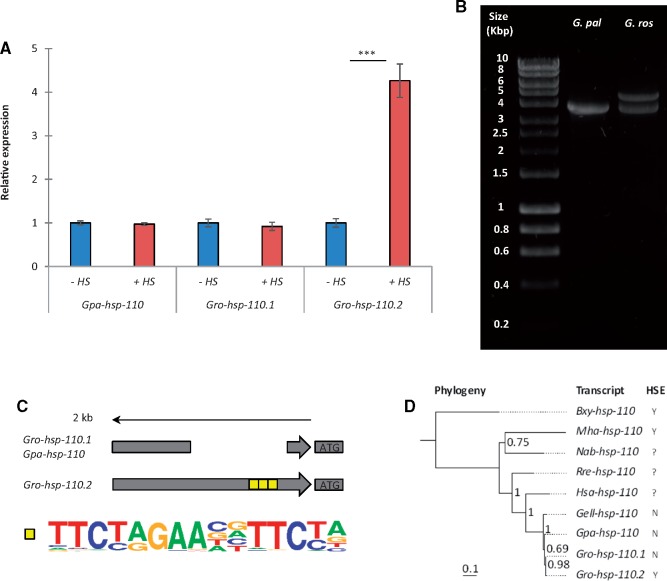
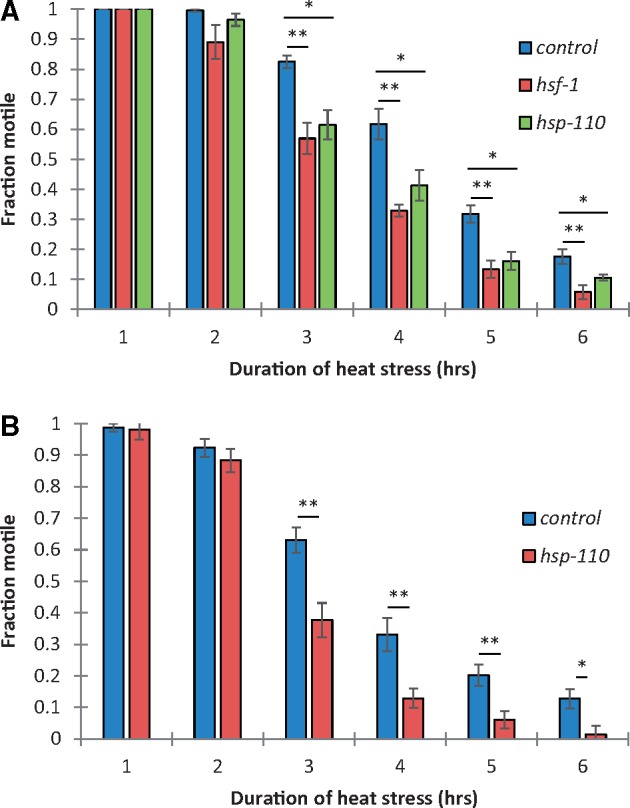
Managing the emergence and spread of crop pests and pathogens is essential for global food security. Understanding how organisms have adapted to their native climate is key to predicting the impact of climate change. The potato cyst nematodes Globodera pallida and G. rostochiensis are economically important plant pathogens that cause yield losses of up to 50% in potato. The two species have different thermal optima that may relate to differences in the altitude of their regions of origin in the Andes. Here, we demonstrate that juveniles of G. pallida are less able to recover from heat stress than those of G. rostochiensis. Genome-wide analysis revealed that while both Globodera species respond to heat stress by induction of various protective heat-inducible genes, G. pallida experiences heat stress at lower temperatures. We use C. elegans as a model to demonstrate the dependence of the heat stress response on expression of Heat Shock Factor-1 (HSF-1). Moreover, we show that hsp-110 is induced by heat stress in G. rostochiensis, but not in the less thermotolerant G. pallida. Sequence analysis revealed that this gene and its promoter was duplicated in G. rostochiensis and acquired thermoregulatory properties. We show that hsp-110 is required for recovery from acute thermal stress in both C. elegans and in G. rostochiensis. Our findings point towards an underlying molecular mechanism that allows the differential expansion of one species relative to another closely related species under current climate change scenarios. Similar mechanisms may be true of other invertebrate species with pest status.
Figures
Similar articles
-
Climate change is predicted to alter the current pest status of Globodera pallida and G. rostochiensis in the United Kingdom.Glob Chang Biol. 2017 Nov;23(11):4497-4507. doi: 10.1111/gcb.13676. Epub 2017 Mar 30. Glob Chang Biol. 2017. PMID: 28261933 Free PMC article.
-
Integrative Morphometric and Molecular Approach to Update the Impact and Distribution of Potato Cyst Nematodes Globodera rostochiensis and Globodera pallida (Tylenchida: Heteroderidae) in Algeria.Pathogens. 2021 Feb 16;10(2):216. doi: 10.3390/pathogens10020216. Pathogens. 2021. PMID: 33669335 Free PMC article.
-
The effect of linalool on second-stage juveniles of the potato cyst nematodes Globodera rostochiensis and G. pallida.J Nematol. 2011 Sep;43(3-4):149-51. J Nematol. 2011. PMID: 23430380 Free PMC article.
-
Molecular variation in the potato cyst nematode, Globodera pallida, in relation to virulence.Commun Agric Appl Biol Sci. 2006;71(3 Pt A):637-8. Commun Agric Appl Biol Sci. 2006. PMID: 17390803 Review.
-
Taxonomy, Morphological and Molecular Identification of the Potato Cyst Nematodes, Globodera pallida and G. rostochiensis.Plants (Basel). 2021 Jan 19;10(1):184. doi: 10.3390/plants10010184. Plants (Basel). 2021. PMID: 33478144 Free PMC article. Review.
Cited by
-
Antioxidant Enzymes and Heat-Shock Protein Genes of Green Peach Aphid (Myzus persicae) Under Short-Time Heat Stress.Front Physiol. 2021 Dec 17;12:805509. doi: 10.3389/fphys.2021.805509. eCollection 2021. Front Physiol. 2021. PMID: 34975546 Free PMC article.
-
Temperature and spinosad-induced modulation of antioxidant enzyme activity and gene expression of adaptive stress-related genes in Ceratitis capitata.Open Vet J. 2025 Jan;15(1):108-117. doi: 10.5455/OVJ.2025.v15.i1.10. Epub 2025 Jan 31. Open Vet J. 2025. PMID: 40092184 Free PMC article. Review.
-
The pervasive impact of global climate change on plant-nematode interaction continuum.Front Plant Sci. 2023 Apr 6;14:1143889. doi: 10.3389/fpls.2023.1143889. eCollection 2023. Front Plant Sci. 2023. PMID: 37089646 Free PMC article. Review.
-
Heat Shock Proteins as Potential Indicators of Induced Stress in Nematodes.Methods Mol Biol. 2024;2756:343-350. doi: 10.1007/978-1-0716-3638-1_13. Methods Mol Biol. 2024. PMID: 38427304
-
Temperature Effects on Expression Levels of hsp Genes in Eggs and Second-Stage Juveniles of Meloidogyne hapla Chitwood, 1949.Int J Mol Sci. 2024 Apr 29;25(9):4867. doi: 10.3390/ijms25094867. Int J Mol Sci. 2024. PMID: 38732085 Free PMC article.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ.. 1990. Basic local alignment search tool. J Mol Biol. 2153: 403–410. - PubMed
-
- Balaban NQ, Merrin J, Chait R, Kowalik L, Leibler S.. 2004. Bacterial persistence as a phenotypic switch. Science 3055690: 1622–1625. - PubMed
-
- Bass C, Field LM.. 2011. Gene amplification and insecticide resistance. Pest Manag Sci. 678: 886–890. - PubMed
-
- Batir JF, Hornbach MJ, Blackwell DD.. 2017. Ten years of measurements and modeling of soil temperature changes and their effects on permafrost in Northwestern Alaska. Glob Planet Change 148:55–71.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
