

De novo variants in GREB1L are associated with non-syndromic inner ear malformations and deafness
- PMID: 29955957
- PMCID: PMC6082420
- DOI: 10.1007/s00439-018-1898-8
De novo variants in GREB1L are associated with non-syndromic inner ear malformations and deafness
Abstract
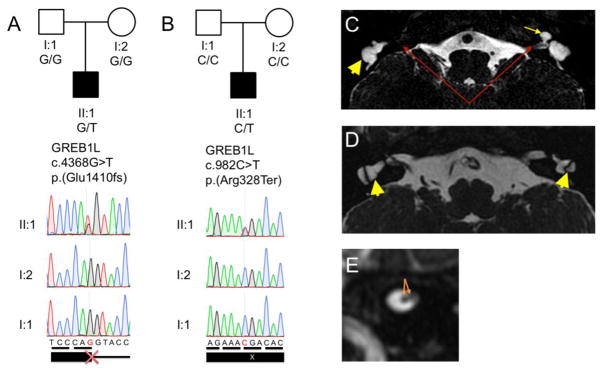
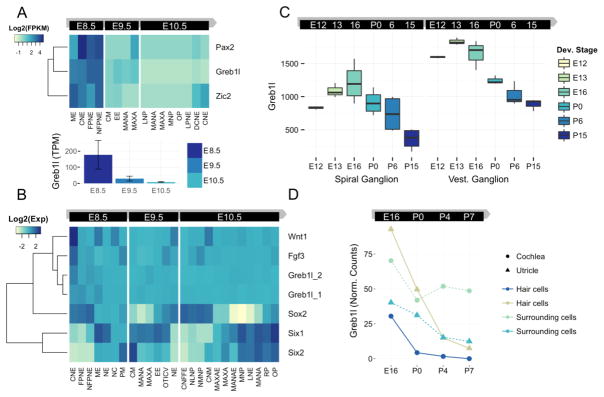
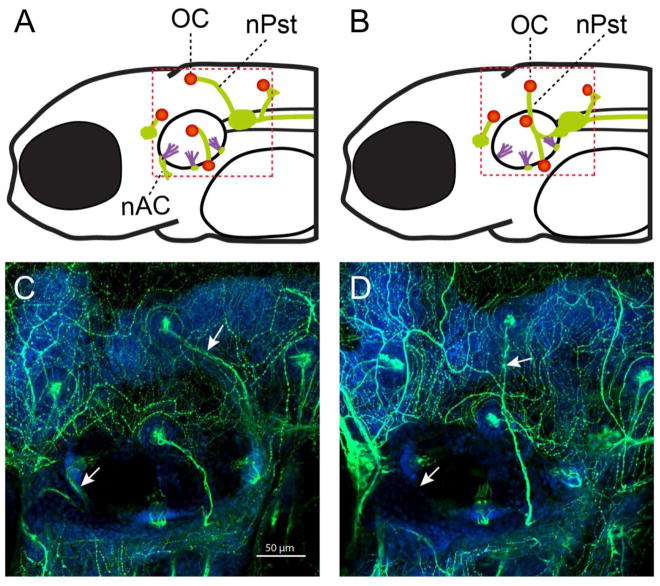
Congenital inner ear malformations affecting both the osseous and membranous labyrinth can have a devastating impact on hearing and language development. With the exception of an enlarged vestibular aqueduct, non-syndromic inner ear malformations are rare, and their underlying molecular biology has thus far remained understudied. To identify molecular factors that might be important in the developing inner ear, we adopted a family-based trio exome sequencing approach in young unrelated subjects with severe inner ear malformations. We identified two previously unreported de novo loss-of-function variants in GREB1L [c.4368G>T;p.(Glu1410fs) and c.982C>T;p.(Arg328*)] in two affected subjects with absent cochleae and eighth cranial nerve malformations. The cochlear aplasia in these affected subjects suggests that a developmental arrest or problem at a very early stage of inner ear development exists, e.g., during the otic pit formation. Craniofacial Greb1l RNA expression peaks in mice during this time frame (E8.5). It also peaks in the developing inner ear during E13-E16, after which it decreases in adulthood. The crucial function of Greb1l in craniofacial development is also evidenced in knockout mice, which develop severe craniofacial abnormalities. In addition, we show that Greb1l-/- zebrafish exhibit a loss of abnormal sensory epithelia innervation. An important role for Greb1l in sensory epithelia innervation development is supported by the eighth cranial nerve deficiencies seen in both affected subjects. In conclusion, we demonstrate that GREB1L is a key player in early inner ear and eighth cranial nerve development. Abnormalities in cochleovestibular anatomy can provide challenges for cochlear implantation. Combining a molecular diagnosis with imaging techniques might aid the development of individually tailored therapeutic interventions in the future.
Figures
Similar articles
-
Autosomal Dominantly Inherited GREB1L Variants in Individuals with Profound Sensorineural Hearing Impairment.Genes (Basel). 2020 Jun 23;11(6):687. doi: 10.3390/genes11060687. Genes (Basel). 2020. PMID: 32585897 Free PMC article.
-
Genes Implicated in Rare Congenital Inner Ear and Cochleovestibular Nerve Malformations.Ear Hear. 2020 Jul/Aug;41(4):983-989. doi: 10.1097/AUD.0000000000000819. Ear Hear. 2020. PMID: 31985533 Free PMC article.
-
Mutations in GREB1L Cause Bilateral Kidney Agenesis in Humans and Mice.Am J Hum Genet. 2017 Nov 2;101(5):803-814. doi: 10.1016/j.ajhg.2017.09.026. Am J Hum Genet. 2017. PMID: 29100091 Free PMC article.
-
Signaling regulating inner ear development: cell fate determination, patterning, morphogenesis, and defects.Congenit Anom (Kyoto). 2015 Feb;55(1):17-25. doi: 10.1111/cga.12072. Congenit Anom (Kyoto). 2015. PMID: 25040109 Review.
-
MicroRNAs and epigenetic regulation in the mammalian inner ear: implications for deafness.Mamm Genome. 2009 Sep-Oct;20(9-10):581-603. doi: 10.1007/s00335-009-9230-5. Epub 2009 Oct 30. Mamm Genome. 2009. PMID: 19876605 Review.
Cited by
-
A novel missense mutation in GREB1L identified in a three-generation family with renal hypodysplasia/aplasia-3.Orphanet J Rare Dis. 2022 Nov 12;17(1):413. doi: 10.1186/s13023-022-02553-w. Orphanet J Rare Dis. 2022. PMID: 36371238 Free PMC article.
-
Diagnostic Yield and Benefits of Whole Exome Sequencing in CAKUT Patients Diagnosed in the First Thousand Days of Life.Kidney Int Rep. 2023 Aug 14;8(11):2439-2457. doi: 10.1016/j.ekir.2023.08.008. eCollection 2023 Nov. Kidney Int Rep. 2023. PMID: 38025229 Free PMC article.
-
Long-range cis-regulatory elements controlling GDF6 expression are essential for ear development.J Clin Invest. 2020 Aug 3;130(8):4213-4217. doi: 10.1172/JCI136951. J Clin Invest. 2020. PMID: 32369452 Free PMC article.
-
A novel autosomal dominant GREB1L variant associated with non-syndromic hearing impairment in Ghana.BMC Med Genomics. 2022 Nov 10;15(1):237. doi: 10.1186/s12920-022-01391-w. BMC Med Genomics. 2022. PMID: 36357908 Free PMC article.
-
A dominant missense variant within LMBR1 related to equine polydactyly.Commun Biol. 2024 Oct 31;7(1):1420. doi: 10.1038/s42003-024-07065-w. Commun Biol. 2024. PMID: 39482424 Free PMC article.
References
-
- Aldrich J, Keats JJ, Liang WS, et al. Abstract 45: Detection of focal somatic copy number variants in whole genome, whole exome, and targeted next-generation sequencing data of tumor/normal pairs. Clin Cancer Res. 2016;22:45–45. doi: 10.1158/1557-3265.PMSCLINGEN15-45. - DOI
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
