

Anti-Correlation between the Dynamics of the Active Site Loop and C-Terminal Tail in Relation to the Homodimer Asymmetry of the Mouse Erythroid 5-Aminolevulinate Synthase
- PMID: 29958424
- PMCID: PMC6073955
- DOI: 10.3390/ijms19071899
Anti-Correlation between the Dynamics of the Active Site Loop and C-Terminal Tail in Relation to the Homodimer Asymmetry of the Mouse Erythroid 5-Aminolevulinate Synthase
Abstract
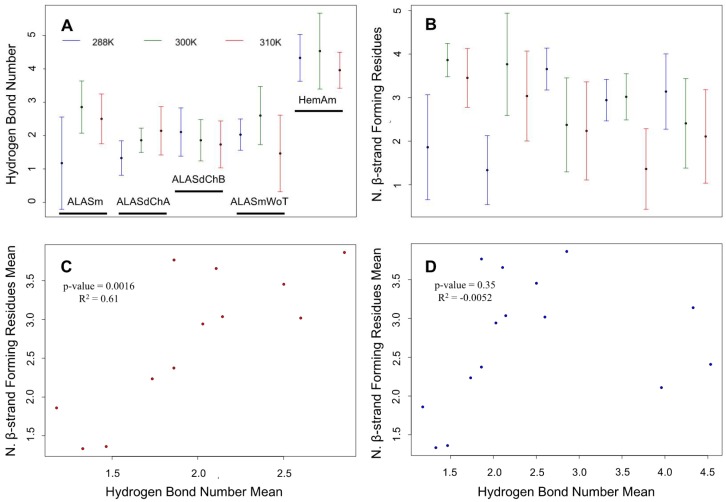
Biosynthesis of heme represents a complex process that involves multiple stages controlled by different enzymes. The first of these proteins is a pyridoxal 5′-phosphate (PLP)-dependent homodimeric enzyme, 5-aminolevulinate synthase (ALAS), that catalyzes the rate-limiting step in heme biosynthesis, the condensation of glycine with succinyl-CoA. Genetic mutations in human erythroid-specific ALAS (ALAS2) are associated with two inherited blood disorders, X-linked sideroblastic anemia (XLSA) and X-linked protoporphyria (XLPP). XLSA is caused by diminished ALAS2 activity leading to decreased ALA and heme syntheses and ultimately ineffective erythropoiesis, whereas XLPP results from “gain-of-function” ALAS2 mutations and consequent overproduction of protoporphyrin IX and increase in Zn2+-protoporphyrin levels. All XLPP-linked mutations affect the intrinsically disordered C-terminal tail of ALAS2. Our earlier molecular dynamics (MD) simulation-based analysis showed that the activity of ALAS2 could be regulated by the conformational flexibility of the active site loop whose structural features and dynamics could be changed due to mutations. We also revealed that the dynamic behavior of the two protomers of the ALAS2 dimer differed. However, how the structural dynamics of ALAS2 active site loop and C-terminal tail dynamics are related to each other and contribute to the homodimer asymmetry remained unanswered questions. In this study, we used bioinformatics and computational biology tools to evaluate the role(s) of the C-terminal tail dynamics in the structure and conformational dynamics of the murine ALAS2 homodimer active site loop. To assess the structural correlation between these two regions, we analyzed their structural displacements and determined their degree of correlation. Here, we report that the dynamics of ALAS2 active site loop is anti-correlated with the dynamics of the C-terminal tail and that this anti-correlation can represent a molecular basis for the functional and dynamic asymmetry of the ALAS2 homodimer.
Keywords: ALAS; anti-correlated dynamics; homodimer asymmetry; intrinsically disordered region; molecular dynamics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
5-Aminolevulinate synthase catalysis: The catcher in heme biosynthesis.Mol Genet Metab. 2019 Nov;128(3):178-189. doi: 10.1016/j.ymgme.2019.06.003. Epub 2019 Jun 13. Mol Genet Metab. 2019. PMID: 31345668 Free PMC article. Review.
-
Elucidating the Role of Human ALAS2 C-terminal Mutations Resulting in Loss of Function and Disease.Biochemistry. 2024 Jul 2;63(13):1636-1646. doi: 10.1021/acs.biochem.4c00066. Epub 2024 Jun 18. Biochemistry. 2024. PMID: 38888931 Free PMC article.
-
Murine erythroid 5-aminolevulinate synthase: Truncation of a disordered N-terminal extension is not detrimental for catalysis.Biochim Biophys Acta. 2016 May;1864(5):441-52. doi: 10.1016/j.bbapap.2016.02.002. Epub 2016 Feb 5. Biochim Biophys Acta. 2016. PMID: 26854603
-
Regulation and tissue-specific expression of δ-aminolevulinic acid synthases in non-syndromic sideroblastic anemias and porphyrias.Mol Genet Metab. 2019 Nov;128(3):190-197. doi: 10.1016/j.ymgme.2019.01.015. Epub 2019 Jan 23. Mol Genet Metab. 2019. PMID: 30737140 Review.
-
The role of genetic testing in accurate diagnosis of X-linked sideroblastic anemia: novel ALAS2 mutations and the impact of X-chromosome inactivation.Sci Rep. 2025 Apr 7;15(1):11843. doi: 10.1038/s41598-025-95590-x. Sci Rep. 2025. PMID: 40195342 Free PMC article.
Cited by
-
The yeast ALA synthase C-terminus positively controls enzyme structure and function.Protein Sci. 2023 Apr;32(4):e4600. doi: 10.1002/pro.4600. Protein Sci. 2023. PMID: 36807942 Free PMC article.
-
Human aminolevulinate synthase structure reveals a eukaryotic-specific autoinhibitory loop regulating substrate binding and product release.Nat Commun. 2020 Jun 4;11(1):2813. doi: 10.1038/s41467-020-16586-x. Nat Commun. 2020. PMID: 32499479 Free PMC article.
-
5-Aminolevulinate synthase catalysis: The catcher in heme biosynthesis.Mol Genet Metab. 2019 Nov;128(3):178-189. doi: 10.1016/j.ymgme.2019.06.003. Epub 2019 Jun 13. Mol Genet Metab. 2019. PMID: 31345668 Free PMC article. Review.
-
An Extended C-Terminus, the Possible Culprit for Differential Regulation of 5-Aminolevulinate Synthase Isoforms.Front Mol Biosci. 2022 Jul 14;9:920668. doi: 10.3389/fmolb.2022.920668. eCollection 2022. Front Mol Biosci. 2022. PMID: 35911972 Free PMC article. Review.
References
-
- Fratz E.J., Stojanovski B.M., Ferreira G.C. Toward Heme: 5-Aminolevulinate Synthase and Initiation of Porphyrin Synthesis. In: Ferreira G.C., Kadish K.M., Smith K.M., Guilard R., editors. Handbook of Porphyrin Science, Heme Biochemistry. Volume 26. World Scientific Publishing Co. Pte. Ltd.; Singapore: 2014. pp. 3–68.
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
