

Inflammatory mediators reduce surface PrPc on human BMVEC resulting in decreased barrier integrity
- PMID: 29959417
- PMCID: PMC6163073
- DOI: 10.1038/s41374-018-0090-z
Inflammatory mediators reduce surface PrPc on human BMVEC resulting in decreased barrier integrity
Abstract
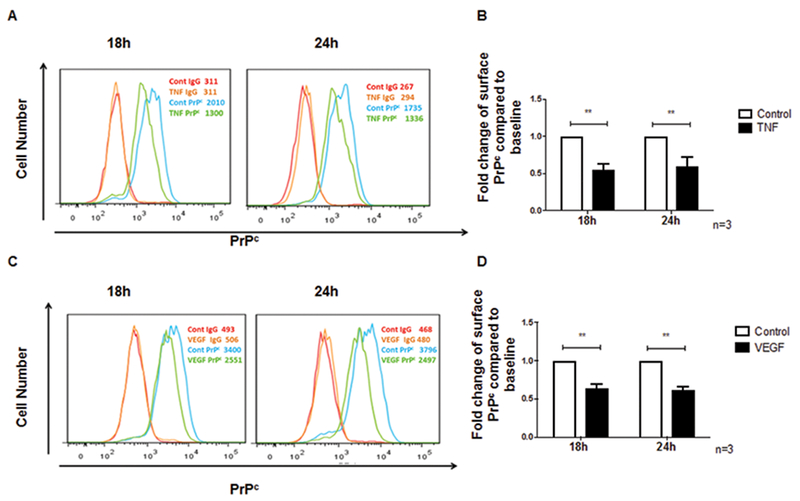
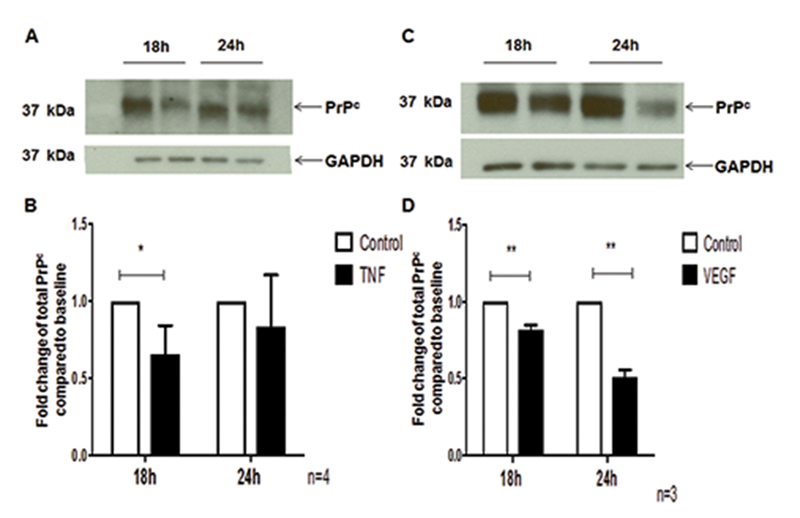
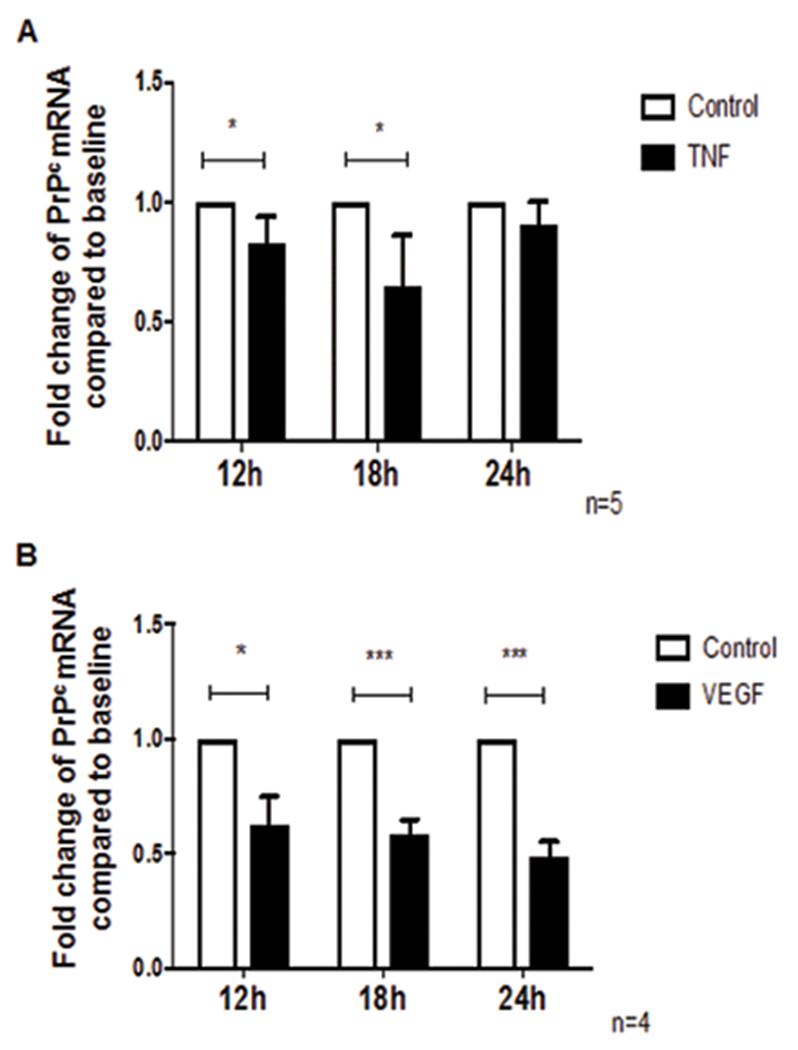
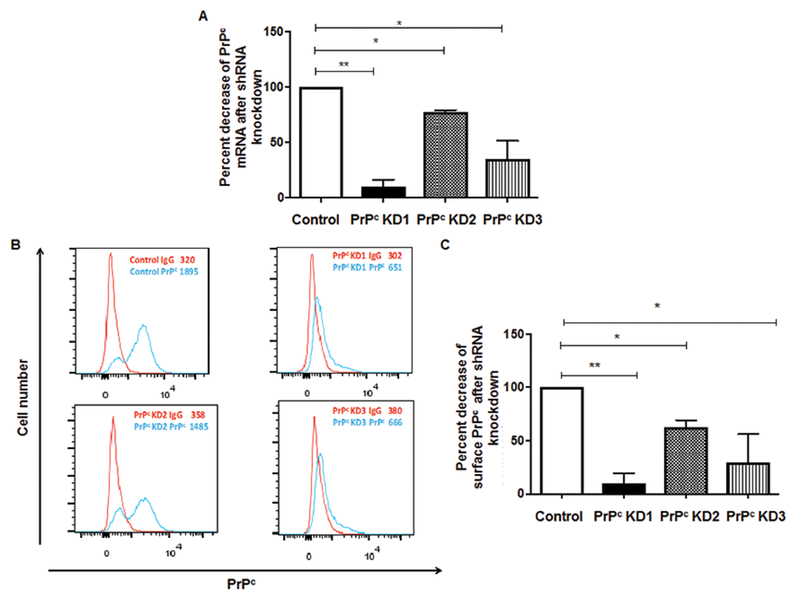
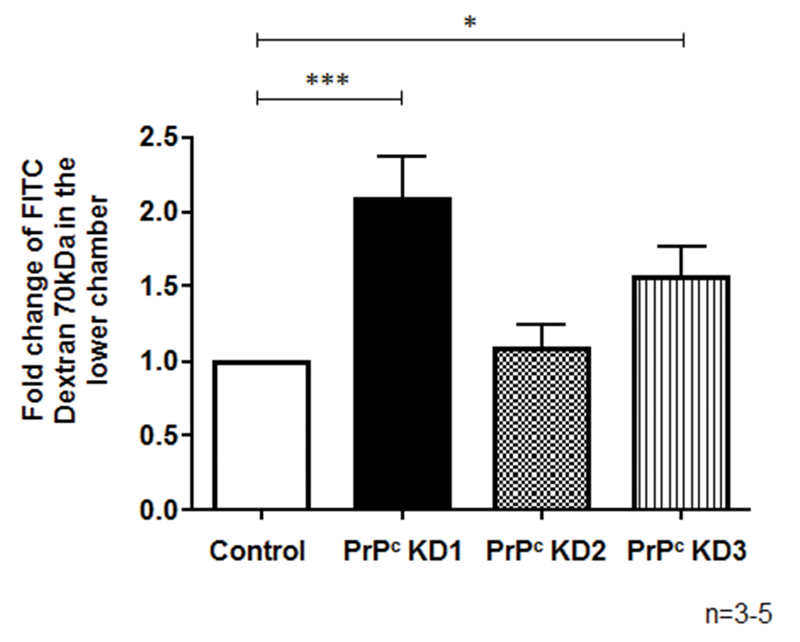
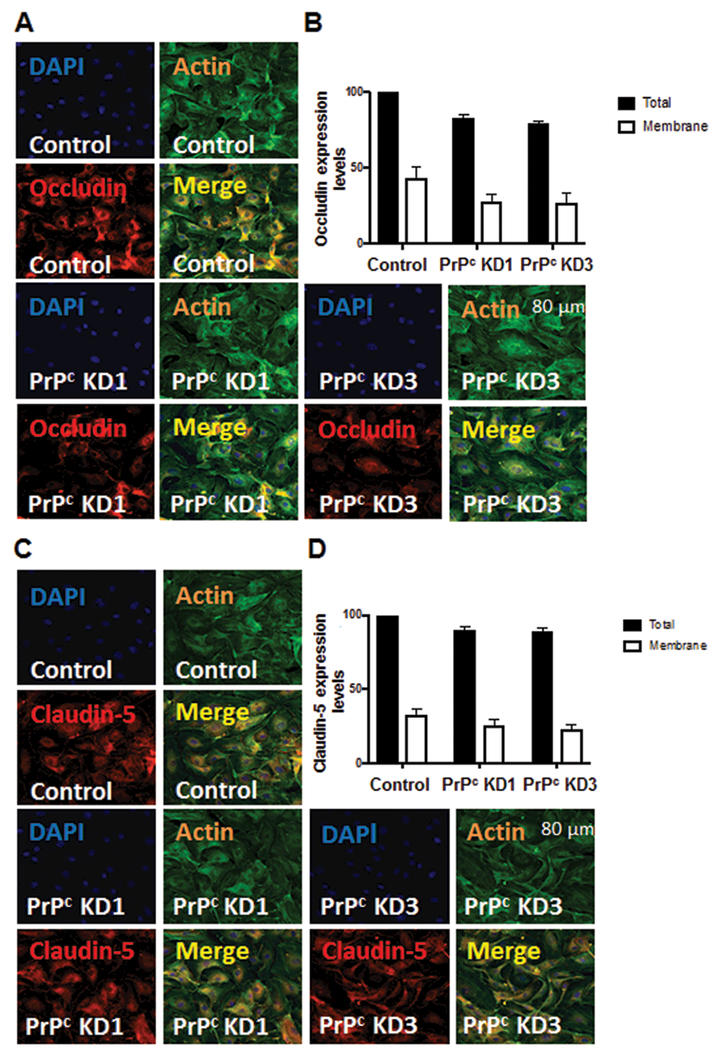
The cellular prion protein (PrPc) is a surface adhesion molecule expressed at junctions of various cell types including brain microvascular endothelial cells (BMVEC) that are important components of the blood-brain barrier (BBB). PrPc is involved in several physiological processes including regulation of epithelial cell barrier function and monocyte migration across BMVEC. BBB dysfunction and disruption are significant events in central nervous system (CNS) inflammatory processes including HIV neuropathogenesis. Tumor necrosis factor (TNF)-α and vascular endothelial growth factor (VEGF) are two inflammatory factors that have been implicated in the processes that affect BBB integrity. To examine the effect of inflammation on PrPc expression in BMVEC, we used these mediators and found that TNF-α and VEGF decrease surface PrPc on primary human BMVEC. We also showed that these factors decrease total PrPc protein as well as mRNA, indicating that they regulate expression of this protein by de novo synthesis. To determine the effect of PrPc loss from the surface of BMVEC on barrier integrity, we used small hairpin RNAs to knockdown PrPc. We found that the absence of PrPc from BMVEC causes increased permeability as determined by a fluorescein isothiocyanate (FITC)-dextran permeability assay. This suggests that cell surface PrPc is essential for endothelial monolayer integrity. To determine the mechanism by which PrPc downregulation leads to increased permeability of an endothelial monolayer, we examined changes in expression and localization of tight junction proteins, occludin and claudin-5, and found that decreased PrPc leads to decreased total and membrane-associated occludin and claudin-5. We propose that an additional mechanism by which inflammatory factors affect endothelial monolayer permeability is by decreasing cell-associated PrPc. This increase in permeability may have subsequent consequences that lead to CNS damage.
Conflict of interest statement
Disclosure/Conflict of Interest
The authors declare no conflict of interest.
Figures
Similar articles
-
miR-27a-3p regulates expression of intercellular junctions at the brain endothelium and controls the endothelial barrier permeability.PLoS One. 2022 Jan 13;17(1):e0262152. doi: 10.1371/journal.pone.0262152. eCollection 2022. PLoS One. 2022. PMID: 35025943 Free PMC article.
-
JAM-A Acts via C/EBP-α to Promote Claudin-5 Expression and Enhance Endothelial Barrier Function.Circ Res. 2020 Sep 25;127(8):1056-1073. doi: 10.1161/CIRCRESAHA.120.316742. Epub 2020 Jul 15. Circ Res. 2020. PMID: 32673519 Free PMC article.
-
Activation of the α7 nicotinic acetylcholine receptor upregulates blood-brain barrier function through increased claudin-5 and occludin expression in rat brain endothelial cells.Neurosci Lett. 2019 Feb 16;694:9-13. doi: 10.1016/j.neulet.2018.11.022. Epub 2018 Nov 16. Neurosci Lett. 2019. PMID: 30452951
-
Blood-brain barrier: structural components and function under physiologic and pathologic conditions.J Neuroimmune Pharmacol. 2006 Sep;1(3):223-36. doi: 10.1007/s11481-006-9025-3. Epub 2006 Jul 6. J Neuroimmune Pharmacol. 2006. PMID: 18040800 Review.
-
The region-selective regulation of endothelial claudin-5 expression and signaling in brain health and disorders.J Cell Physiol. 2021 Oct;236(10):7134-7143. doi: 10.1002/jcp.30357. Epub 2021 Mar 10. J Cell Physiol. 2021. PMID: 33694168 Review.
Cited by
-
In vitro modeling of blood-brain barrier and interface functions in neuroimmune communication.Fluids Barriers CNS. 2020 Mar 30;17(1):26. doi: 10.1186/s12987-020-00187-3. Fluids Barriers CNS. 2020. PMID: 32228633 Free PMC article. Review.
-
Blood-Brain Barrier Dysfunction Amplifies the Development of Neuroinflammation: Understanding of Cellular Events in Brain Microvascular Endothelial Cells for Prevention and Treatment of BBB Dysfunction.Front Cell Neurosci. 2021 Sep 13;15:661838. doi: 10.3389/fncel.2021.661838. eCollection 2021. Front Cell Neurosci. 2021. PMID: 34588955 Free PMC article. Review.
-
Physiology of Cellular Prion Proteins in Reproduction.Dev Reprod. 2024 Jun;28(2):29-36. doi: 10.12717/DR.2024.28.2.29. Epub 2024 Jun 30. Dev Reprod. 2024. PMID: 39055100 Free PMC article.
-
Prion Protein at the Leading Edge: Its Role in Cell Motility.Int J Mol Sci. 2020 Sep 12;21(18):6677. doi: 10.3390/ijms21186677. Int J Mol Sci. 2020. PMID: 32932634 Free PMC article. Review.
-
Show Me Your Friends and I Tell You Who You Are: The Many Facets of Prion Protein in Stroke.Cells. 2020 Jul 2;9(7):1609. doi: 10.3390/cells9071609. Cells. 2020. PMID: 32630841 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
