

Four Key Steps Control Glycolytic Flux in Mammalian Cells
- PMID: 29960885
- PMCID: PMC6062487
- DOI: 10.1016/j.cels.2018.06.003
Four Key Steps Control Glycolytic Flux in Mammalian Cells
Abstract
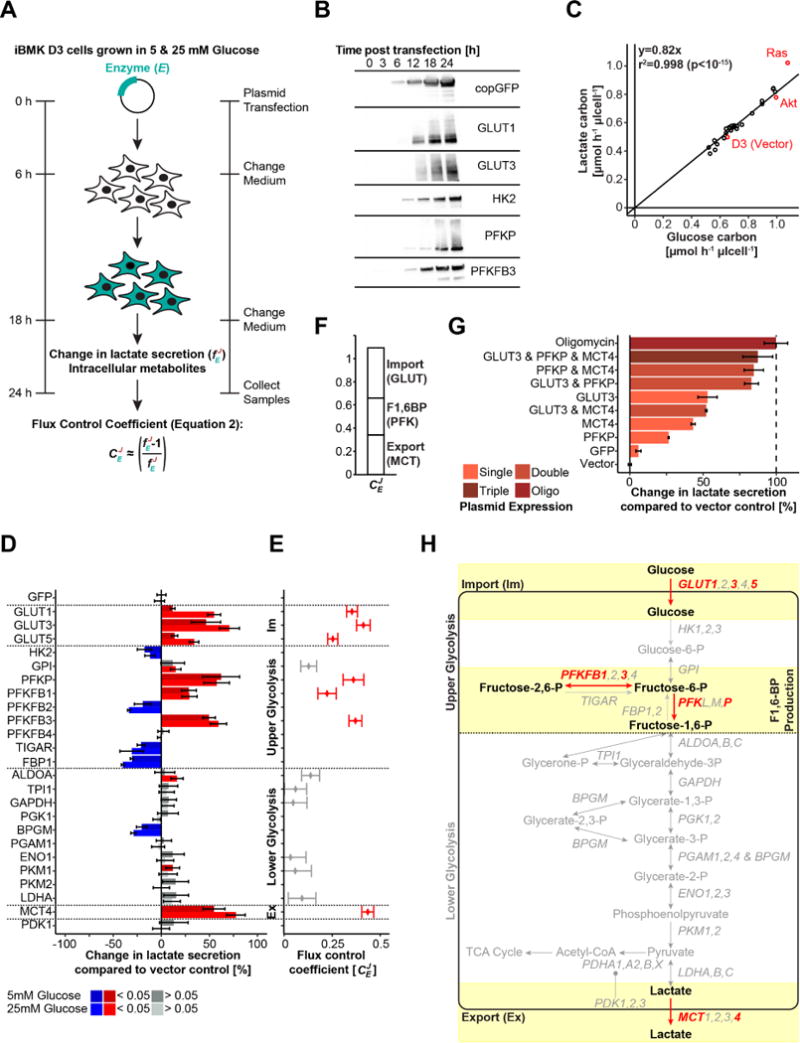
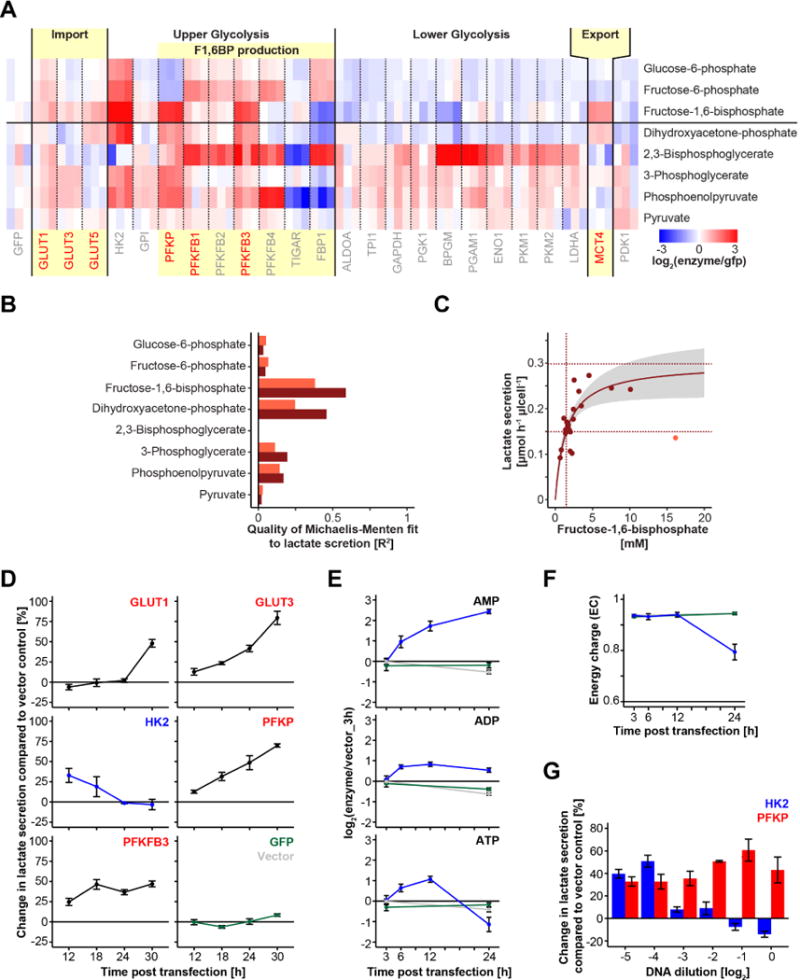
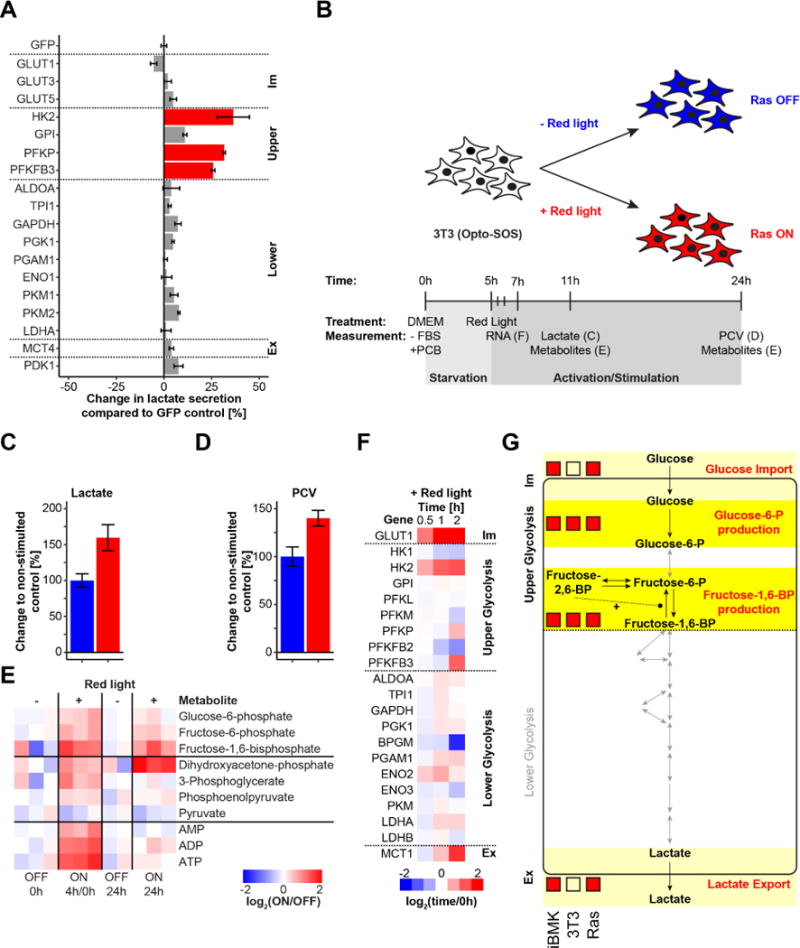
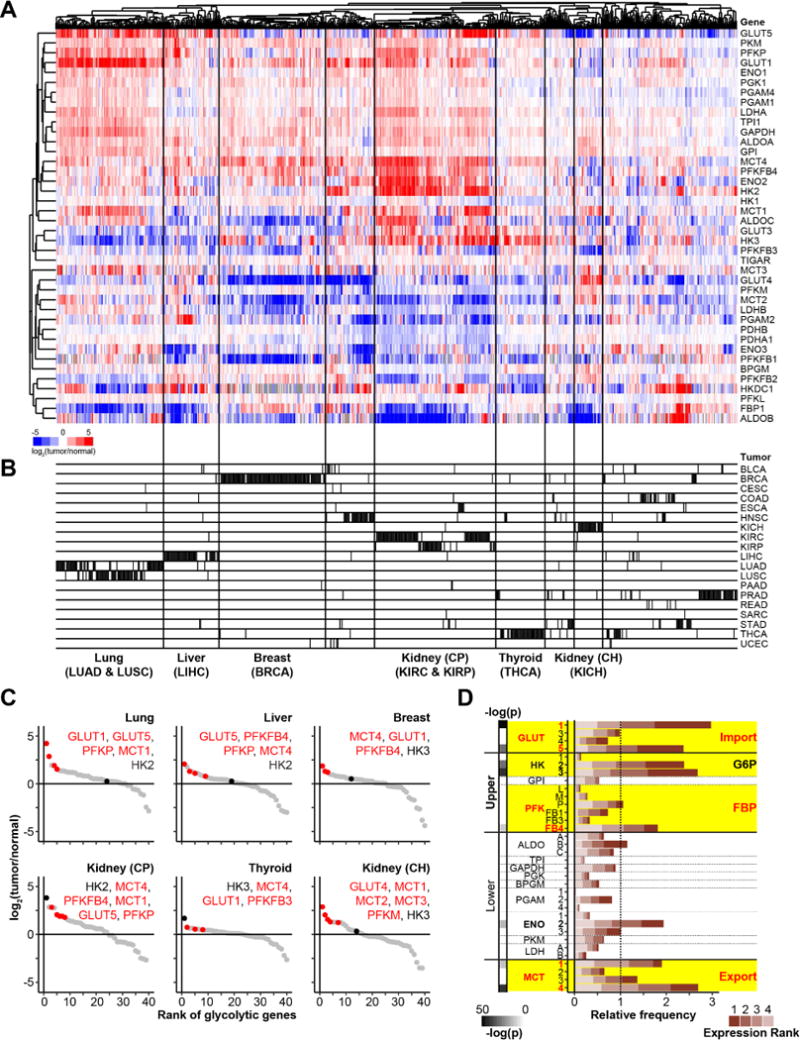
Altered glycolysis is a hallmark of diseases including diabetes and cancer. Despite intensive study of the contributions of individual glycolytic enzymes, systems-level analyses of flux control through glycolysis remain limited. Here, we overexpress in two mammalian cell lines the individual enzymes catalyzing each of the 12 steps linking extracellular glucose to excreted lactate, and find substantial flux control at four steps: glucose import, hexokinase, phosphofructokinase, and lactate export (and not at any steps of lower glycolysis). The four flux-controlling steps are specifically upregulated by the Ras oncogene: optogenetic Ras activation rapidly induces the transcription of isozymes catalyzing these four steps and enhances glycolysis. At least one isozyme catalyzing each of these four steps is consistently elevated in human tumors. Thus, in the studied contexts, flux control in glycolysis is concentrated in four key enzymatic steps. Upregulation of these steps in tumors likely underlies the Warburg effect.
Keywords: GLUT; MCT1; PFK; PFKFB; flux control; glycolysis; metabolic control analysis; metabolism; metabolomics; optogenetics.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Abdelmalek MF, Lazo M, Horska A, Bonekamp S, Lipkin EW, Balasubramanyam A, Bantle JP, Johnson RJ, Diehl AM, Clark JM, et al. Higher dietary fructose is associated with impaired hepatic adenosine triphosphate homeostasis in obese individuals with type 2 diabetes. Hepatology. 2012;56:952–960. - PMC - PubMed
-
- Abu-Amero KK, Alzahrani AS, Zou M, Shi Y. High frequency of somatic mitochondrial DNA mutations in human thyroid carcinomas and complex I respiratory defect in thyroid cancer cell lines. Oncogene. 2005;24:1455–1460. - PubMed
-
- Van Aelst L, Hohmann S, Bulaya B, de Koning W, Sierkstra L, Neves MJ, Luyten K, Alijo R, Ramos J, Coccetti P. Molecular cloning of a gene involved in glucose sensing in the yeast Saccharomyces cerevisiae. Mol Microbiol. 1993;8:927–943. - PubMed
-
- Bensaad K, Tsuruta A, Selak MA, Vidal MNC, Nakano K, Bartrons R, Gottlieb E, Vousden KH. TIGAR, a p53-inducible regulator of glycolysis and apoptosis. Cell. 2006;126:107–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
