

Mouse but not zebrafish requires retinoic acid for control of neuromesodermal progenitors and body axis extension
- PMID: 29964026
- PMCID: PMC6064660
- DOI: 10.1016/j.ydbio.2018.06.019
Mouse but not zebrafish requires retinoic acid for control of neuromesodermal progenitors and body axis extension
Abstract
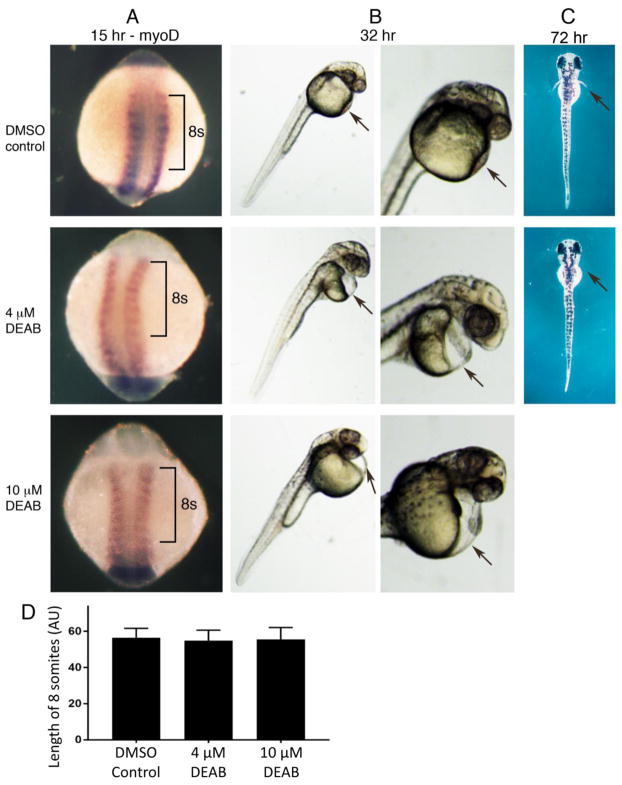
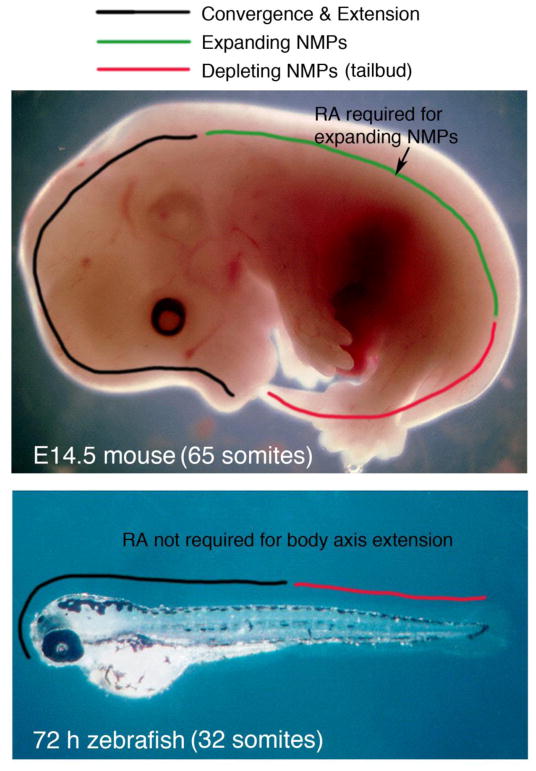
In mouse, retinoic acid (RA) is required for the early phase of body axis extension controlled by a population of neuromesodermal progenitors (NMPs) in the trunk called expanding-NMPs, but not for the later phase of body axis extension controlled by a population of NMPs in the tail called depleting-NMPs. Recent observations suggest that zebrafish utilize depleting-NMPs but not expanding-NMPs for body axis extension. In zebrafish, a role for RA in body axis extension was not supported by previous studies on aldh1a2 (raldh2) mutants lacking RA synthesis. Here, by treating zebrafish embryos with an RA synthesis inhibitor, we also found that body axis extension and somitogenesis was not perturbed, although loss of pectoral fin and cardiac edema were observed consistent with previous studies. The conclusion that zebrafish diverges from mouse in not requiring RA for body axis extension is consistent with zebrafish lacking early expanding-NMPs to generate the trunk. We suggest that RA control of body axis extension was added to higher vertebrates during evolution of expanding-NMPs.
Keywords: Body axis extension; NMPs; Neuromesodermal progenitors; Retinoic acid; Somitogenesis.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Amin S, Neijts R, Simmini S, van Rooijen C, Tan SC, Kester L, van Oudenaarden A, Creyghton MP, Deschamps J. Cdx and T Brachyury Co-activate Growth Signaling in the Embryonic Axial Progenitor Niche. Cell Reports. 2016;17:3165–3177. - PubMed
-
- Begemann G, Schilling TF, Rauch GJ, Geisler R, Ingham PW. The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development. 2001;128:3081–3094. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
