

Differential expression and function of CAIX and CAXII in breast cancer: A comparison between tumorgraft models and cells
- PMID: 29965974
- PMCID: PMC6028082
- DOI: 10.1371/journal.pone.0199476
Differential expression and function of CAIX and CAXII in breast cancer: A comparison between tumorgraft models and cells
Expression of concern in
-
Expression of Concern: Differential expression and function of CAIX and CAXII in breast cancer: A comparison between tumorgraft models and cells.PLoS One. 2025 Jul 3;20(7):e0327518. doi: 10.1371/journal.pone.0327518. eCollection 2025. PLoS One. 2025. PMID: 40608685 Free PMC article. No abstract available.
Abstract
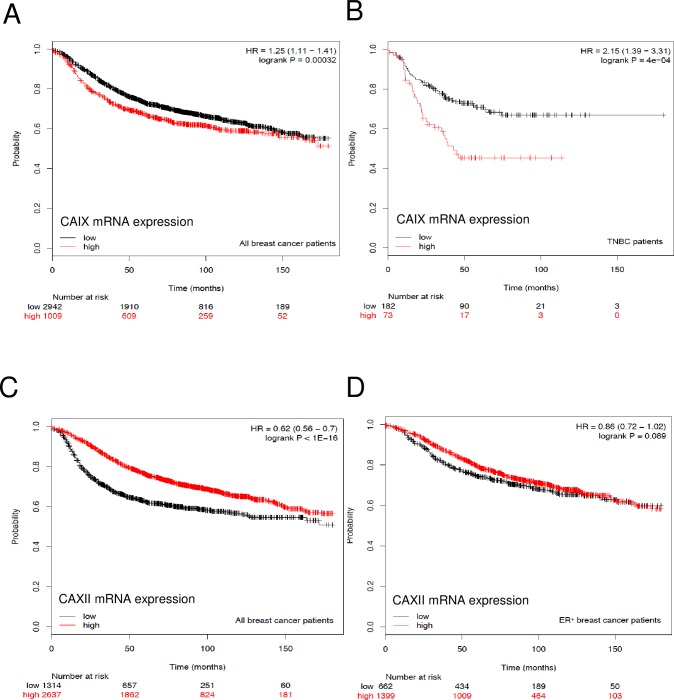
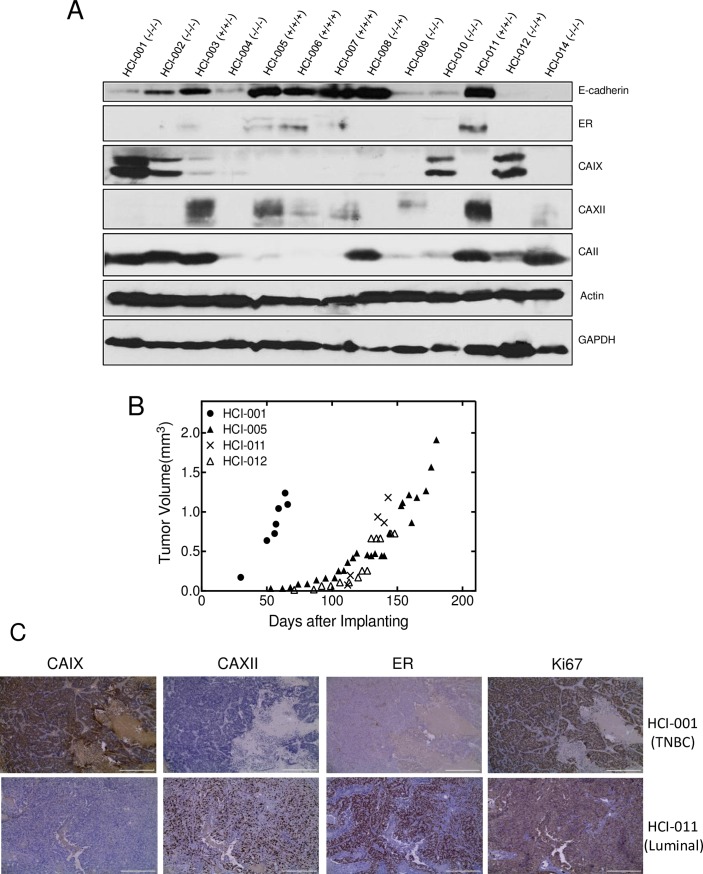
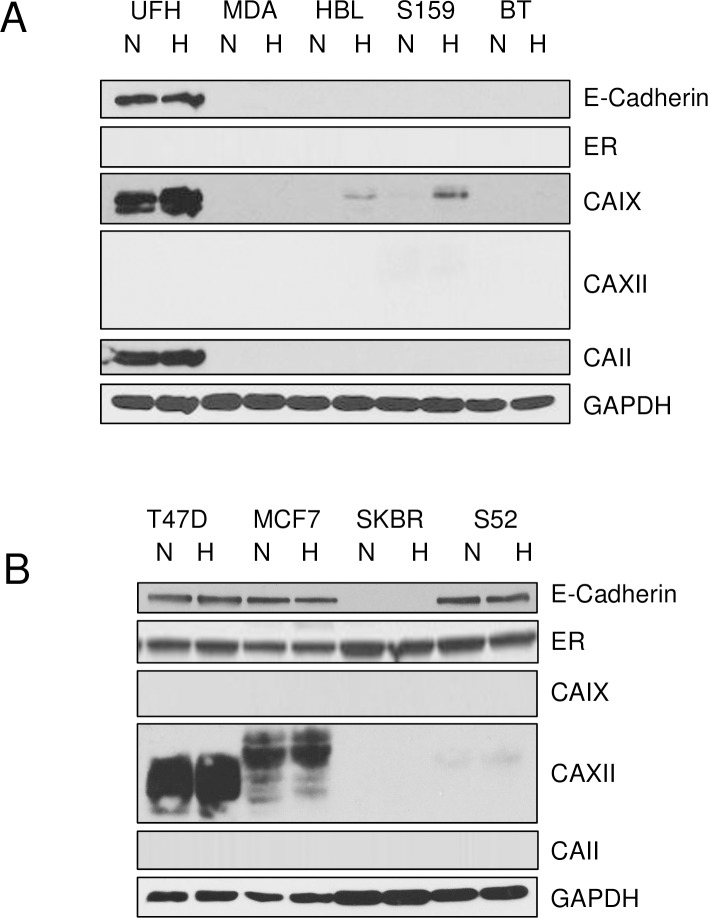
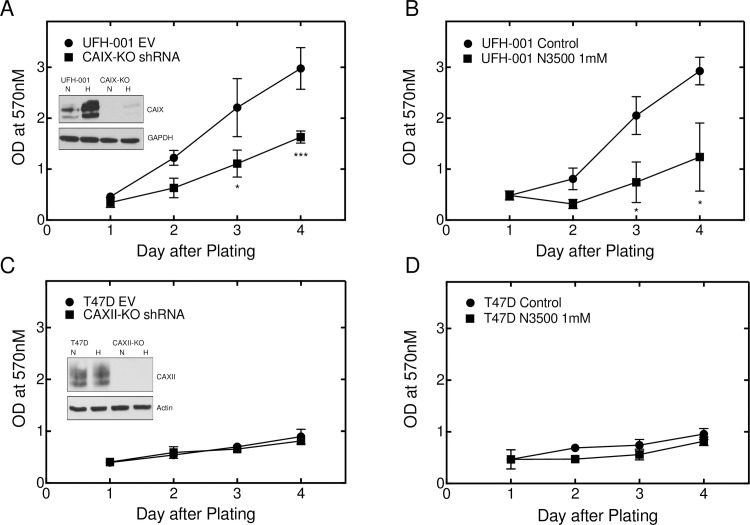
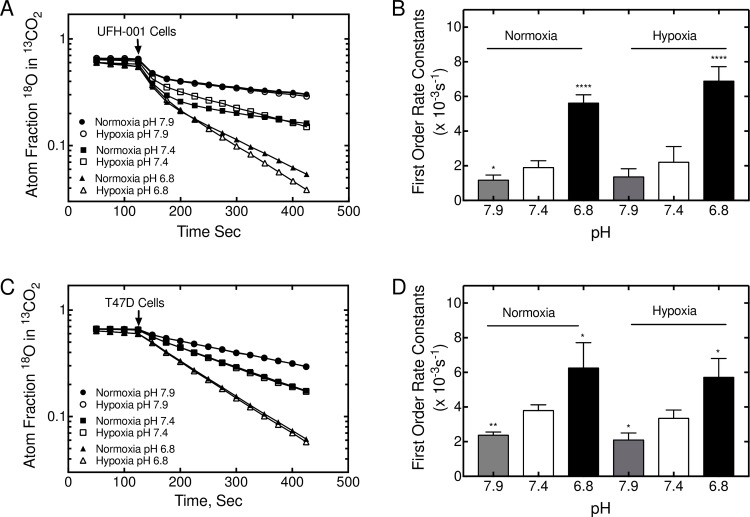
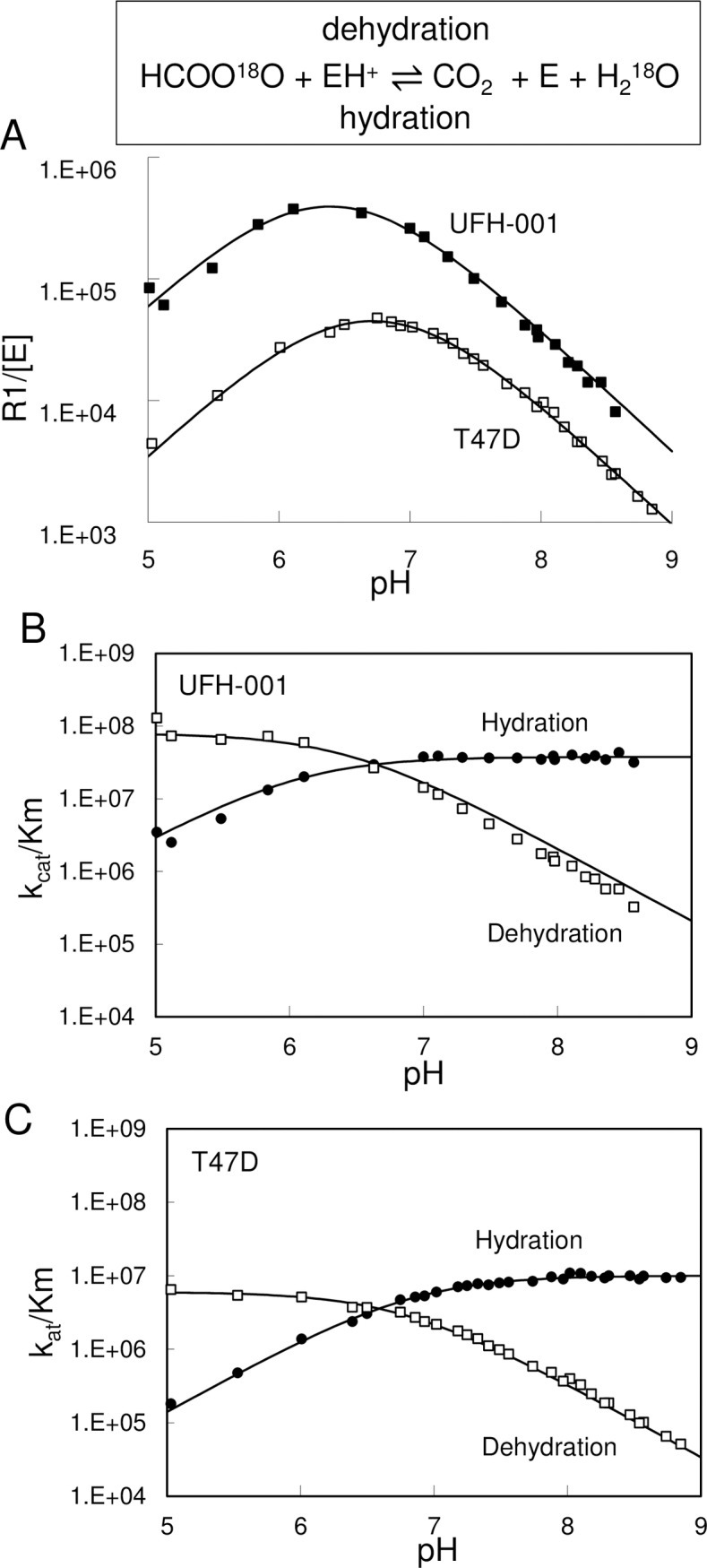
Carbonic anhydrase IX (CAIX) and XII (CAXII) are transmembrane proteins that are associated with cancer progression. We have previously described the catalytic properties of CAIX in MDA-MB-231 breast cancer cells, a line of cells that were derived from a patient with triple negative breast cancer. We chose this line because CAIX expression in breast cancer is a marker of hypoxia and a prognosticator for reduced survival. However, CAXII expression is associated with better survival statistics than those patients with low CAXII expression. Yet CAIX and CAXII have similar catalytic activities. Here we compare the potential roles of CAIX and CAXII in the context of TNBC and estrogen receptor (ER)-positive breast cancer. In tumor graft models, we show that CAIX and CAXII exhibit distinct expression patterns and non-overlapping. We find the same pattern across a panel of TNBC and luminal breast cancer cell lines. This affords an opportunity to compare directly CAIX and CAXII function. Our data suggest that CAIX expression is associated with growth potentiation in the tumor graft model and in a TNBC line using knockdown strategies and blocking activity with an impermeant sulfonamide inhibitor, N-3500. CAXII was not associated with growth potentiation. The catalytic activities of both CAIX and CAXII were sensitive to inhibition by N-3500 and activated at low pH. However, pH titration of activity in membrane ghosts revealed significant differences in the catalytic efficiency and pKa values. These features provide evidence that CAIX is a more efficient enzyme than CAXII at low pH and that CAIX shifts the equilibrium between CO2 and bicarbonate in favor of CO2 production by consuming protons. This suggests that in the acidic microenvironment of tumors, CAIX plays a role in stabilizing pH at a value that favors cancer cell survival.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
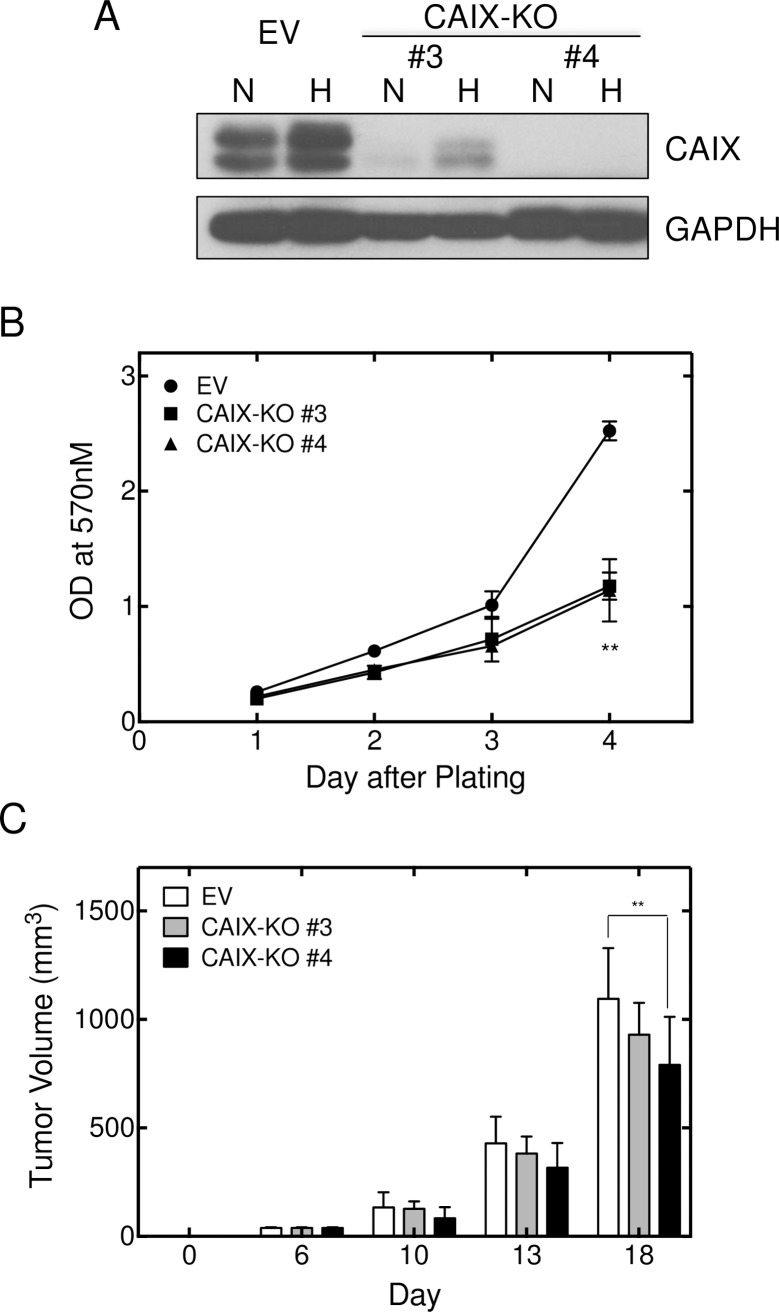
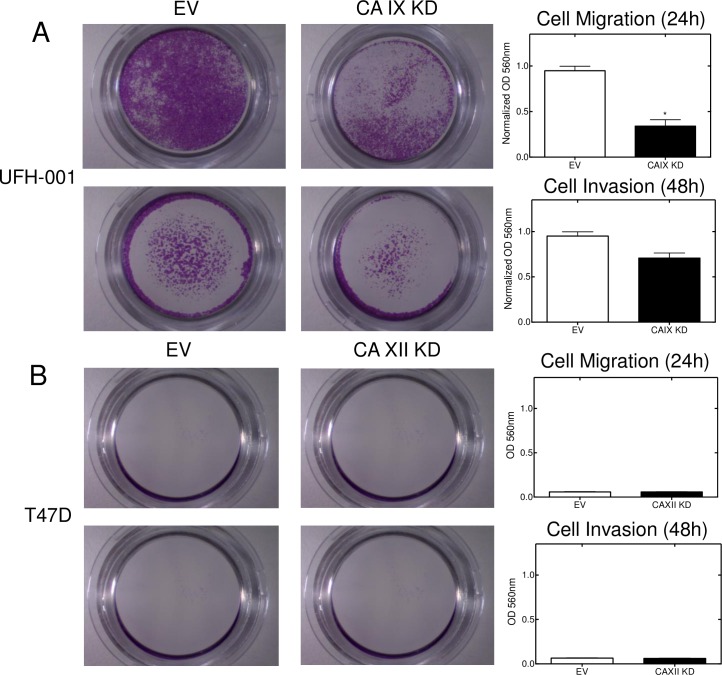
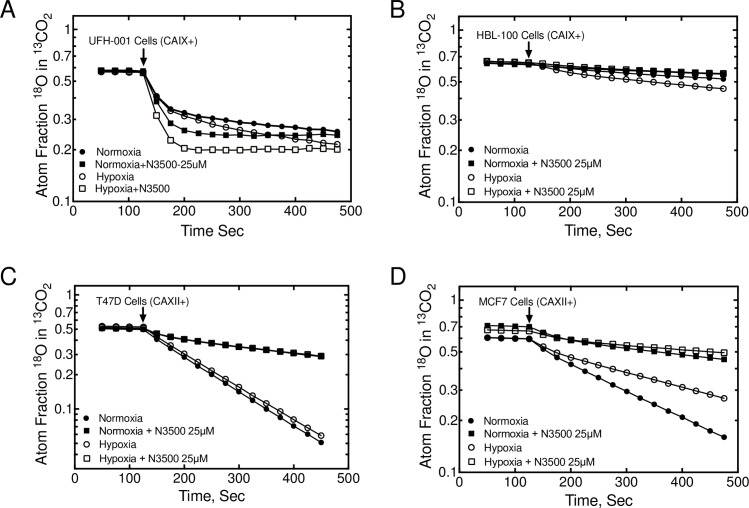
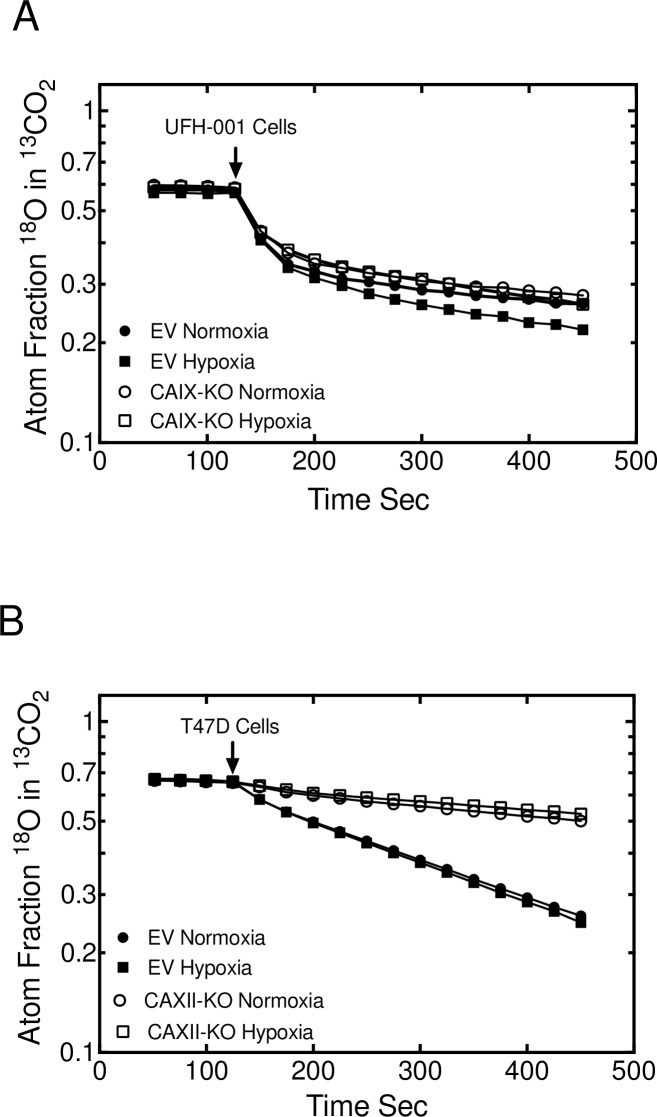
References
-
- Schwickert G, Walenta S, Sundfor K, Rofstad EK, Mueller-Klieser W. Correlation of High Lactate Levels in Human Cervical Cancer with Incidence of Metastasis. Cancer Res. 1995;55:4757–9. - PubMed
-
- Helmlinger G, Sckell A, Dellian M, Forbes NS, Jain RK. Acid Production in Glycolysis-impaired Tumors Provides New Insights into Tumor Metabolism. Clin Canc Res 2002;8:1284–91. - PubMed
-
- Frost SC. Physiological functions of the alpha class of carbonic anhydrases. Subcell Biochem. 2014;75:9–30. doi: 10.1007/978-94-007-7359-2_2 . - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
