

Inhibition of integrin αVβ6 changes fibril thickness of stromal collagen in experimental carcinomas
- PMID: 29966518
- PMCID: PMC6027735
- DOI: 10.1186/s12964-018-0249-7
Inhibition of integrin αVβ6 changes fibril thickness of stromal collagen in experimental carcinomas
Abstract
Background: Chemotherapeutic efficacy can be improved by targeting the structure and function of the extracellular matrix (ECM) in the carcinomal stroma. This can be accomplished by e.g. inhibiting TGF-β1 and -β3 or treating with Imatinib, which results in scarcer collagen fibril structure in xenografted human KAT-4/HT29 (KAT-4) colon adenocarcinoma.
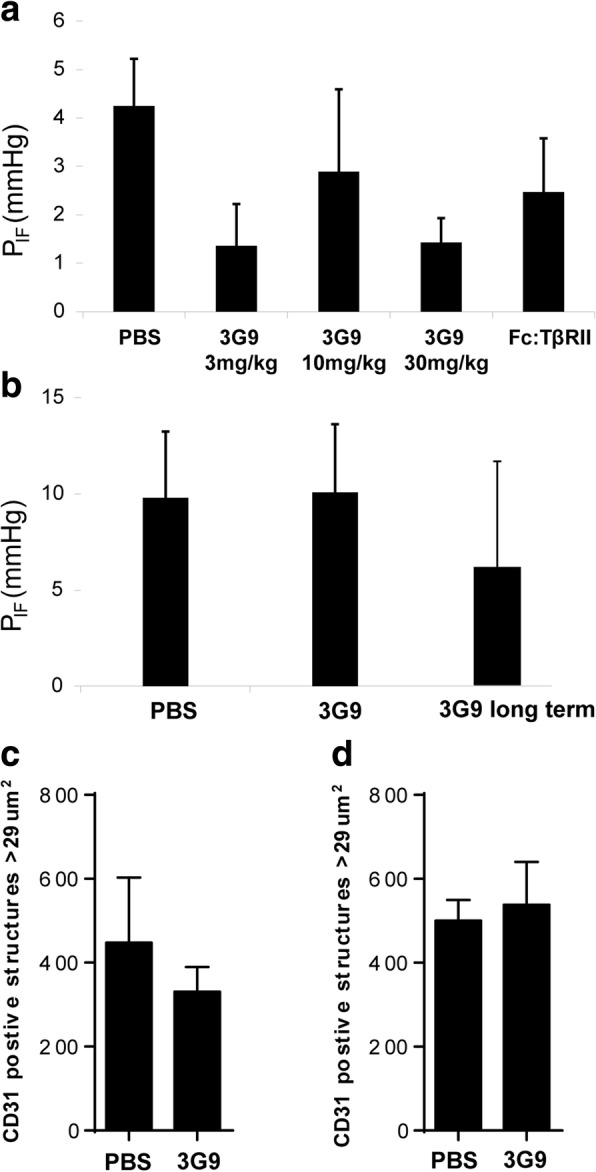
Methods: The potential role of αVβ6 integrin-mediated activation of latent TGF-β was studied in cultured KAT-4 and Capan-2 human ductal pancreatic carcinoma cells as well as in xenograft carcinoma generated by these cells. The monoclonal αVβ6 integrin-specific monoclonal antibody 3G9 was used to inhibit the αVβ6 integrin activity.
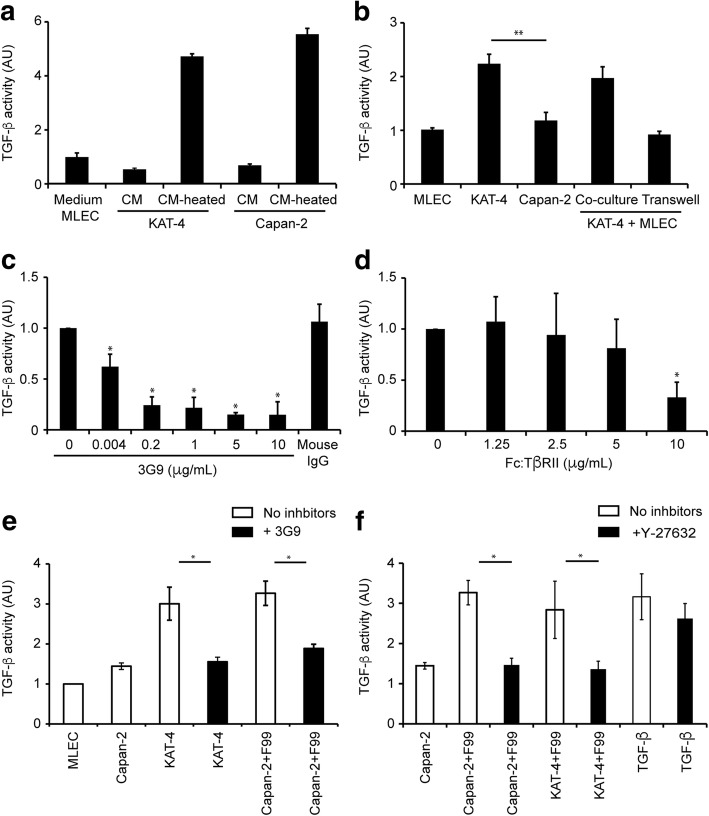
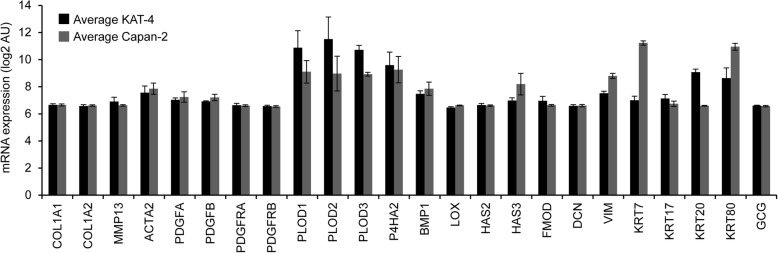
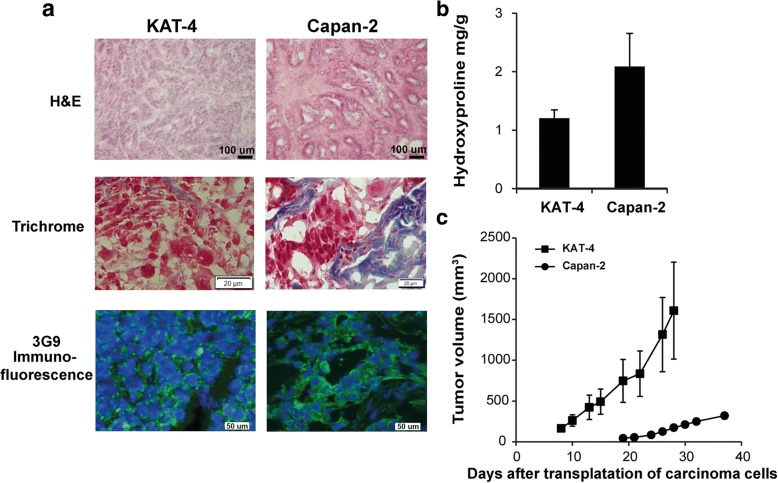
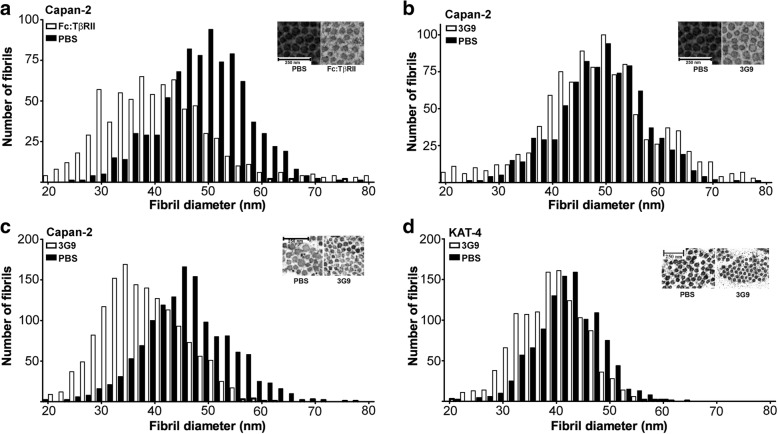
Results: Both KAT-4 and Capan-2 cells expressed the αVβ6 integrin but only KAT-4 cells could utilize this integrin to activate latent TGF-β in vitro. Only when Capan-2 cells were co-cultured with human F99 fibroblasts was the integrin activation mechanism triggered, suggesting a more complex, fibroblast-dependent, activation pathway. In nude mice, a 10-day treatment with 3G9 reduced collagen fibril thickness and interstitial fluid pressure in KAT-4 but not in the more desmoplastic Capan-2 tumors that, to achieve a similar effect, required a prolonged 3G9 treatment. In contrast, a 10-day direct inhibition of TGF-β1 and -β3 reduced collagen fibril thickness in both tumor models.
Conclusion: Our data demonstrate that the αVβ6-directed activation of latent TGF-β plays a pivotal role in modulating the stromal collagen network in carcinoma, but that the sensitivity to αVβ6 inhibition depends on the simultaneous presence of alternative paths for latent TGF-β activation and the extent of desmoplasia.
Conflict of interest statement
Ethics approval
All animal experiments were performed at the animal facilities of the National Veterinary Institute and Lund University, Sweden, in accordance with approval by the ethical committees for animal experiments in Uppsala and Malmö/Lund, Sweden. The number of animals was minimized to comply with guidelines from the Ethical Committee and EU legislation on the use of laboratory animals.
Consent for publication
All authors approved the final version of the manuscript and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All persons designated as authors qualify for authorship, and all those who qualify for authorship are listed.
Competing interests
Three authors (LAK, PHW, SMV) were employees of Biogen at the time the work was completed. One of the authors (PHW) still is an employee at Biogen. None of the other authors report any potential competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
A human monoclonal antibody 264RAD targeting αvβ6 integrin reduces tumour growth and metastasis, and modulates key biomarkers in vivo.Oncogene. 2013 Sep 12;32(37):4406-16. doi: 10.1038/onc.2012.460. Epub 2012 Oct 29. Oncogene. 2013. PMID: 23108397
-
Pro-migratory and TGF-β-activating functions of αvβ6 integrin in pancreatic cancer are differentially regulated via an Eps8-dependent GTPase switch.J Pathol. 2017 Sep;243(1):37-50. doi: 10.1002/path.4923. Epub 2017 Aug 7. J Pathol. 2017. PMID: 28608476 Free PMC article.
-
Influenza promotes collagen deposition via αvβ6 integrin-mediated transforming growth factor β activation.J Biol Chem. 2014 Dec 19;289(51):35246-63. doi: 10.1074/jbc.M114.582262. Epub 2014 Oct 22. J Biol Chem. 2014. PMID: 25339175 Free PMC article.
-
Epithelial-mesenchymal interactions in fibrosis and repair. Transforming growth factor-β activation by epithelial cells and fibroblasts.Ann Am Thorac Soc. 2015 Mar;12 Suppl 1(Suppl 1):S21-3. doi: 10.1513/AnnalsATS.201406-245MG. Ann Am Thorac Soc. 2015. PMID: 25830829 Free PMC article. Review.
-
Integrin αvβ6: Structure, function and role in health and disease.Int J Biochem Cell Biol. 2018 Jun;99:186-196. doi: 10.1016/j.biocel.2018.04.013. Epub 2018 Apr 18. Int J Biochem Cell Biol. 2018. PMID: 29678785 Review.
Cited by
-
Targeting TGF-β signal transduction for fibrosis and cancer therapy.Mol Cancer. 2022 Apr 23;21(1):104. doi: 10.1186/s12943-022-01569-x. Mol Cancer. 2022. PMID: 35461253 Free PMC article. Review.
-
Identification of integrin β6 gene promoter and analysis of its transcription regulation in colon cancer cells.World J Gastrointest Oncol. 2020 May 15;12(5):526-534. doi: 10.4251/wjgo.v12.i5.526. World J Gastrointest Oncol. 2020. PMID: 32461784 Free PMC article.
-
ITGB6 promotes pancreatic fibrosis and aggravates the malignant process of pancreatic cancer via JAK2/STAT3 signaling pathway.Naunyn Schmiedebergs Arch Pharmacol. 2024 Aug;397(8):6093-6106. doi: 10.1007/s00210-024-03003-z. Epub 2024 Feb 28. Naunyn Schmiedebergs Arch Pharmacol. 2024. PMID: 38418753
-
Integrin Alpha v Beta 6 (αvβ6) and Its Implications in Cancer Treatment.Int J Mol Sci. 2022 Oct 15;23(20):12346. doi: 10.3390/ijms232012346. Int J Mol Sci. 2022. PMID: 36293202 Free PMC article. Review.
-
The Role of the Transforming Growth Factor-β Signaling Pathway in Gastrointestinal Cancers.Biomolecules. 2023 Oct 19;13(10):1551. doi: 10.3390/biom13101551. Biomolecules. 2023. PMID: 37892233 Free PMC article. Review.
References
-
- Nagy JA, Brown LF, Senger DR, Lanir N, Van de Water L, Dvorak AM, Dvorak HF. Pathogenesis of tumor stroma generation: a critical role for leaky blood vessels and fibrin deposition. Biochim Biophys Acta. 1989;948(3):305–326. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
