

HEAT INDUCIBLE LIPASE1 Remodels Chloroplastic Monogalactosyldiacylglycerol by Liberating α-Linolenic Acid in Arabidopsis Leaves under Heat Stress
- PMID: 29967047
- PMCID: PMC6139690
- DOI: 10.1105/tpc.18.00347
HEAT INDUCIBLE LIPASE1 Remodels Chloroplastic Monogalactosyldiacylglycerol by Liberating α-Linolenic Acid in Arabidopsis Leaves under Heat Stress
Abstract
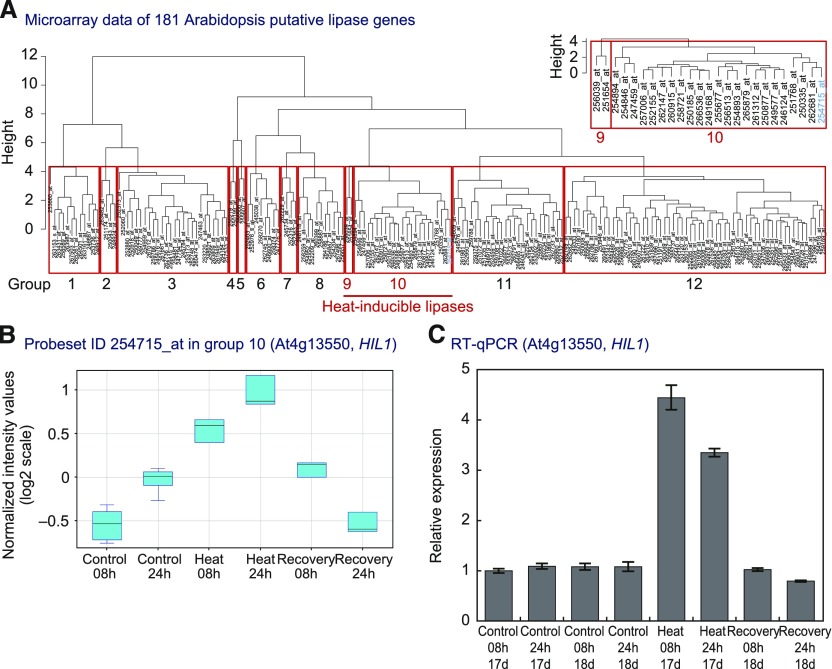
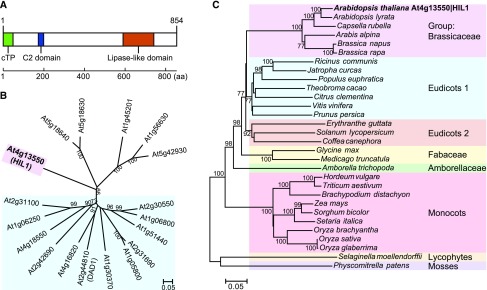
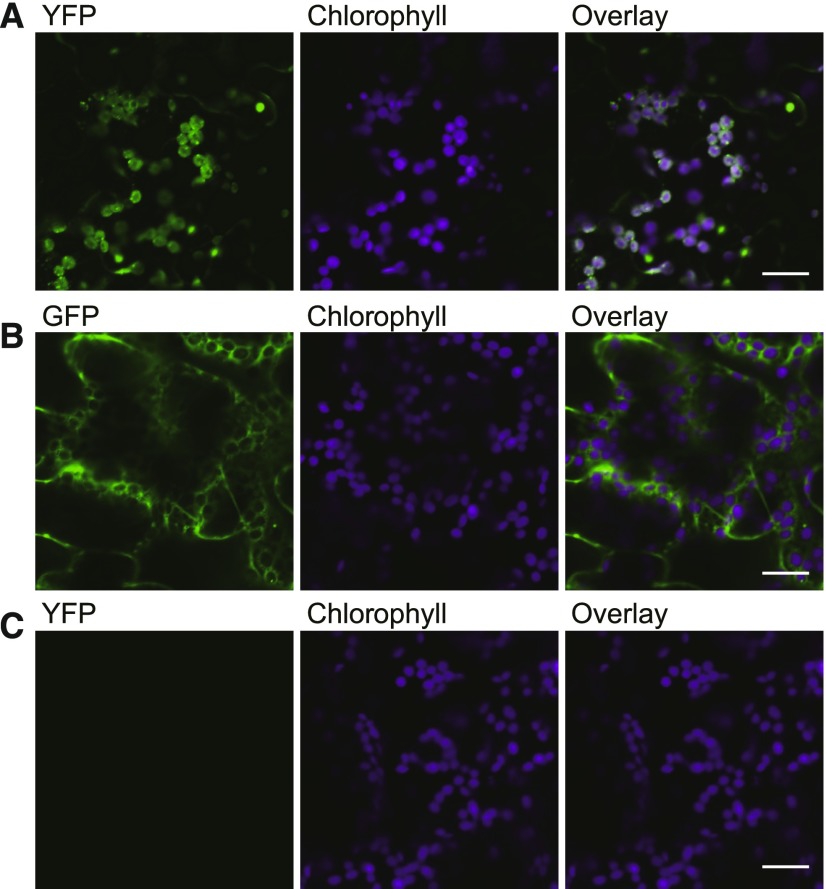
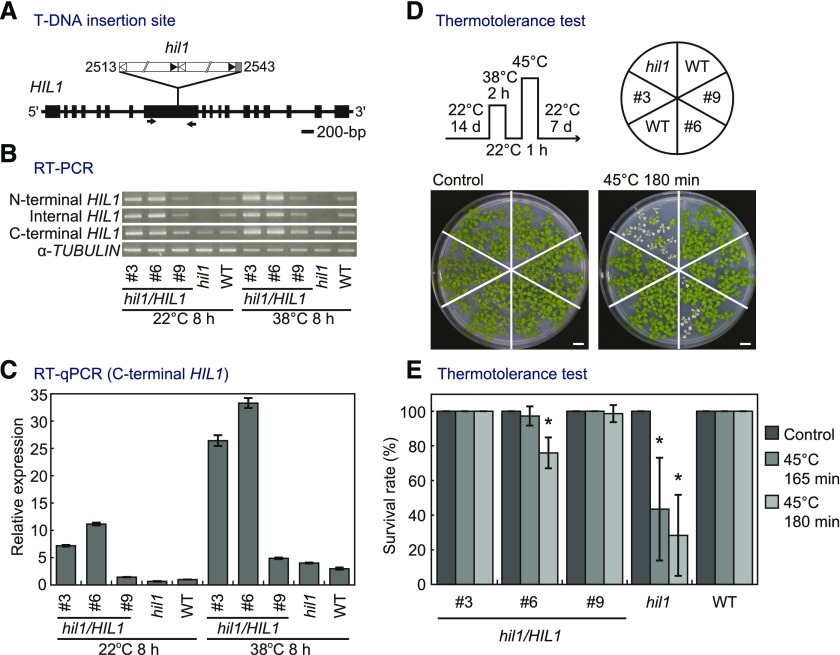
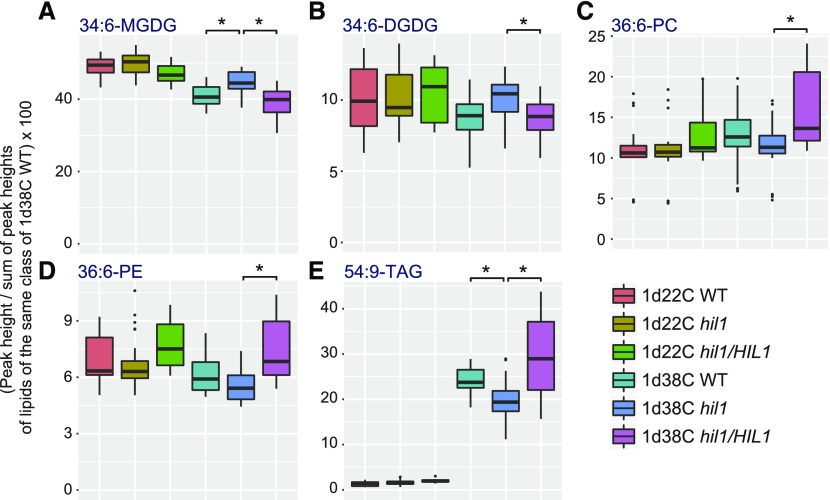
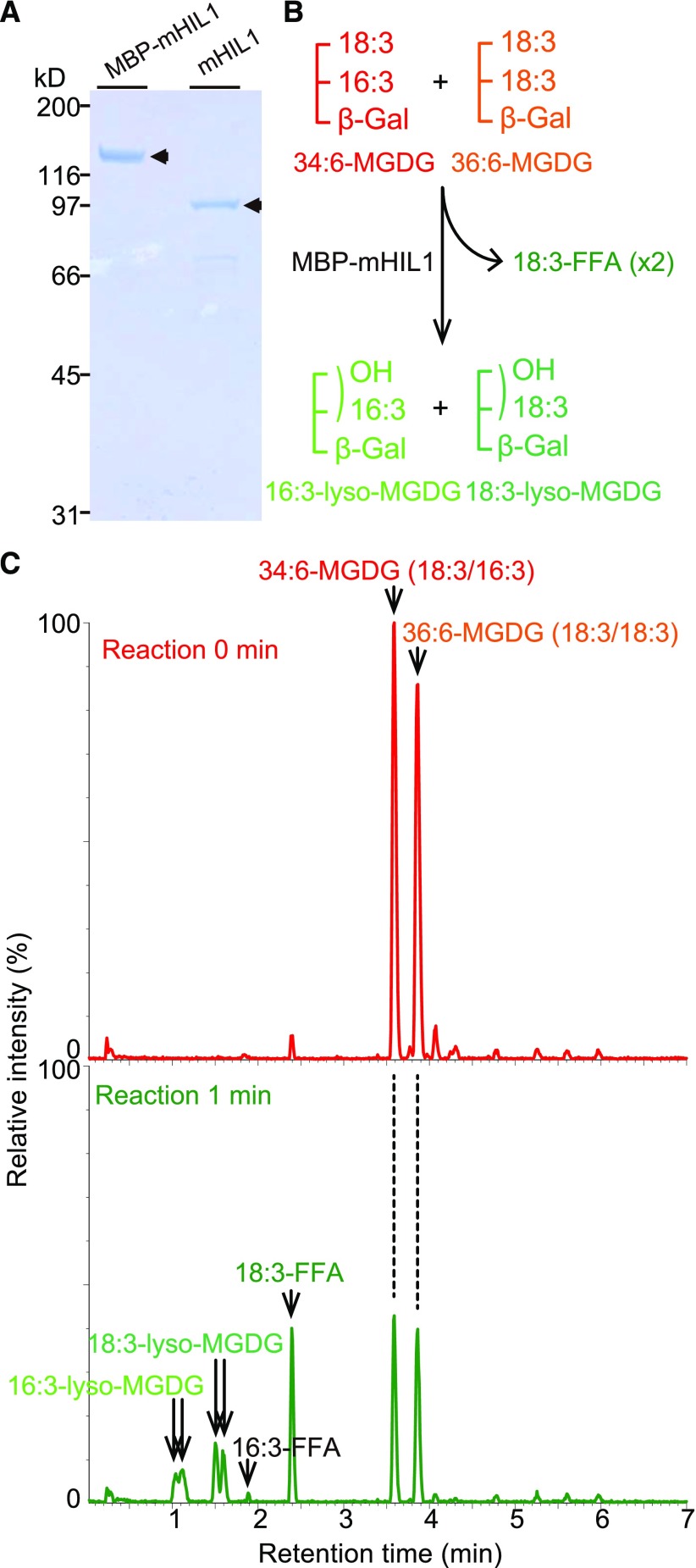
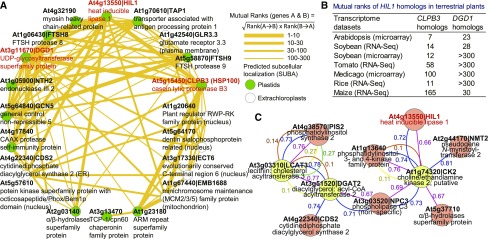
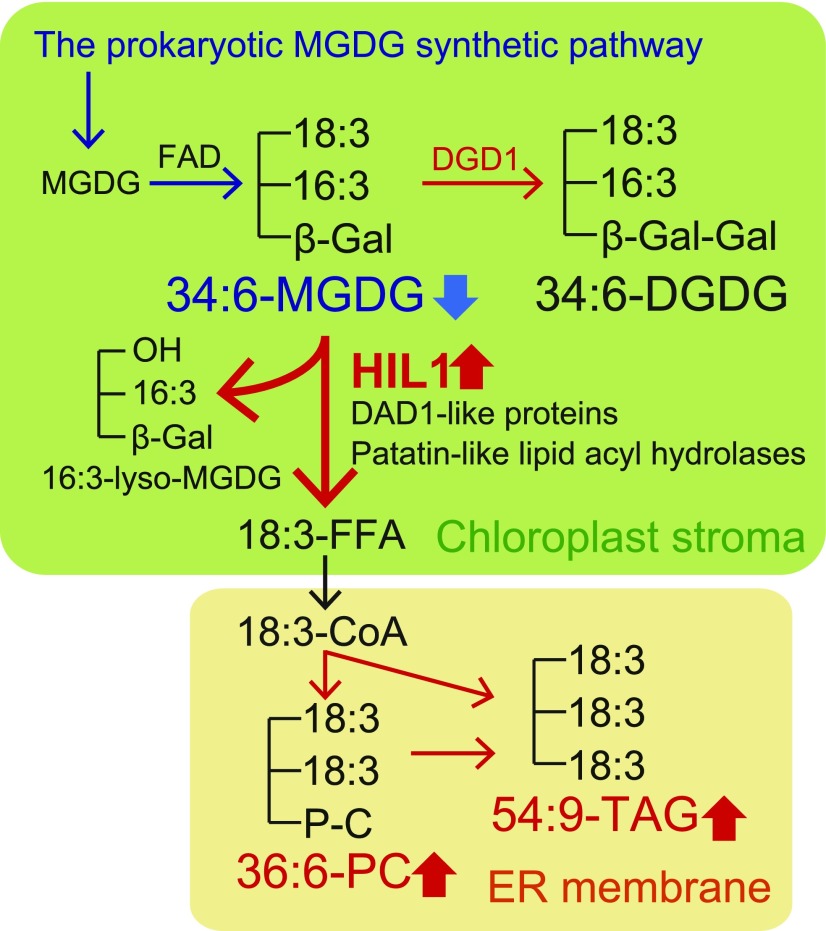
Under heat stress, polyunsaturated acyl groups, such as α-linolenate (18:3) and hexadecatrienoate (16:3), are removed from chloroplastic glycerolipids in various plant species. Here, we showed that a lipase designated HEAT INDUCIBLE LIPASE1 (HIL1) induces the catabolism of monogalactosyldiacylglycerol (MGDG) under heat stress in Arabidopsis thaliana leaves. Using thermotolerance tests, a T-DNA insertion mutant with disrupted HIL1 was shown to have a heat stress-sensitive phenotype. Lipidomic analysis indicated that the decrease of 34:6-MGDG under heat stress was partially impaired in the hil1 mutant. Concomitantly, the heat-induced increment of 54:9-triacylglycerol in the hil1 mutant was 18% lower than that in the wild-type plants. Recombinant HIL1 protein digested MGDG to produce 18:3-free fatty acid (18:3-FFA), but not 18:0- and 16:0-FFAs. A transient assay using fluorescent fusion proteins confirmed chloroplastic localization of HIL1. Transcriptome coexpression network analysis using public databases demonstrated that the HIL1 homolog expression levels in various terrestrial plants are tightly associated with chloroplastic heat stress responses. Thus, HIL1 encodes a chloroplastic MGDG lipase that releases 18:3-FFA in the first committed step of 34:6 (18:3/16:3)-containing galactolipid turnover, suggesting that HIL1 has an important role in the lipid remodeling process induced by heat stress in plants.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures

Comment in
-
Heat Trims the Fat: HIL1 Functions in Lipid Homeostasis.Plant Cell. 2018 Aug;30(8):1670-1671. doi: 10.1105/tpc.18.00593. Epub 2018 Aug 10. Plant Cell. 2018. PMID: 30099380 Free PMC article. No abstract available.
References
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657. - PubMed
-
- Bao X., Focke M., Pollard M., Ohlrogge J. (2000). Understanding in vivo carbon precursor supply for fatty acid synthesis in leaf tissue. Plant J. 22: 39–50. - PubMed
-
- Bligh E.G., Dyer W.J. (1959). A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37: 911–917. - PubMed
-
- Chen J., Burke J.J., Xin Z., Xu C., Velten J. (2006). Characterization of the Arabidopsis thermosensitive mutant atts02 reveals an important role for galactolipids in thermotolerance. Plant Cell Environ. 29: 1437–1448. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
