

BrlR from Pseudomonas aeruginosa is a receptor for both cyclic di-GMP and pyocyanin
- PMID: 29967320
- PMCID: PMC6028453
- DOI: 10.1038/s41467-018-05004-y
BrlR from Pseudomonas aeruginosa is a receptor for both cyclic di-GMP and pyocyanin
Abstract
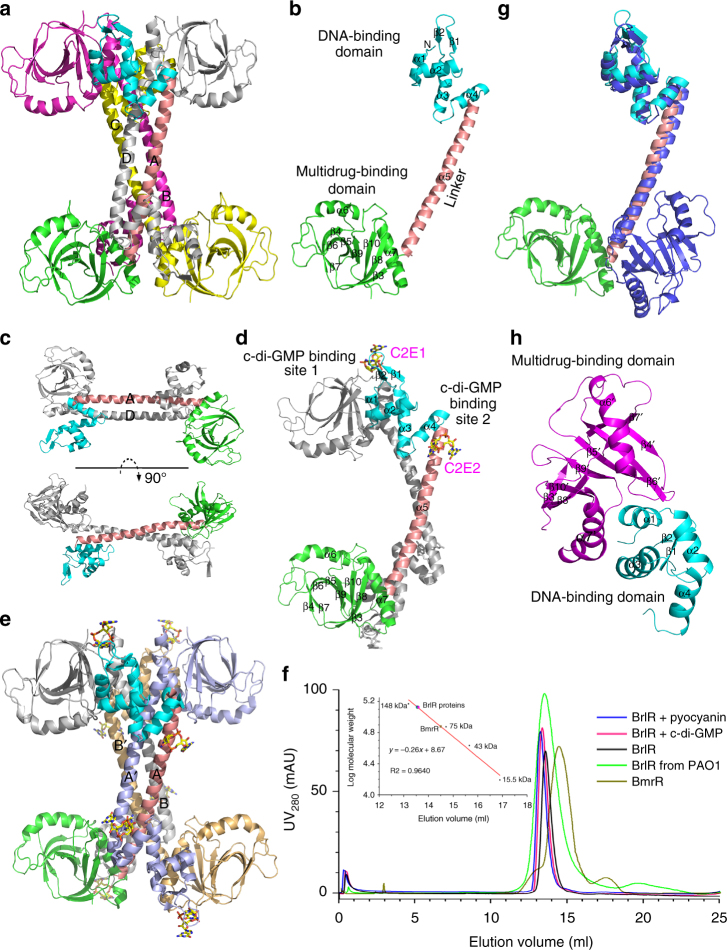
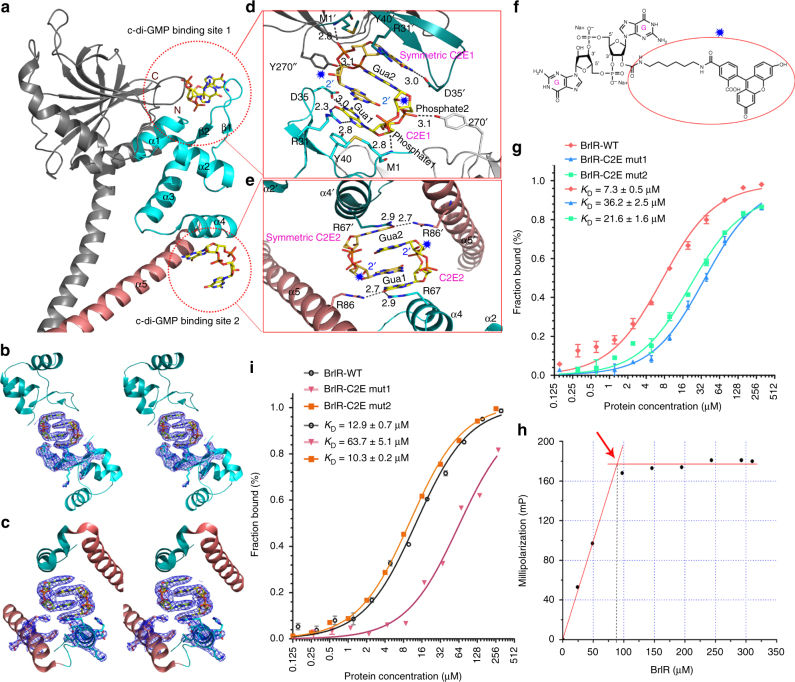
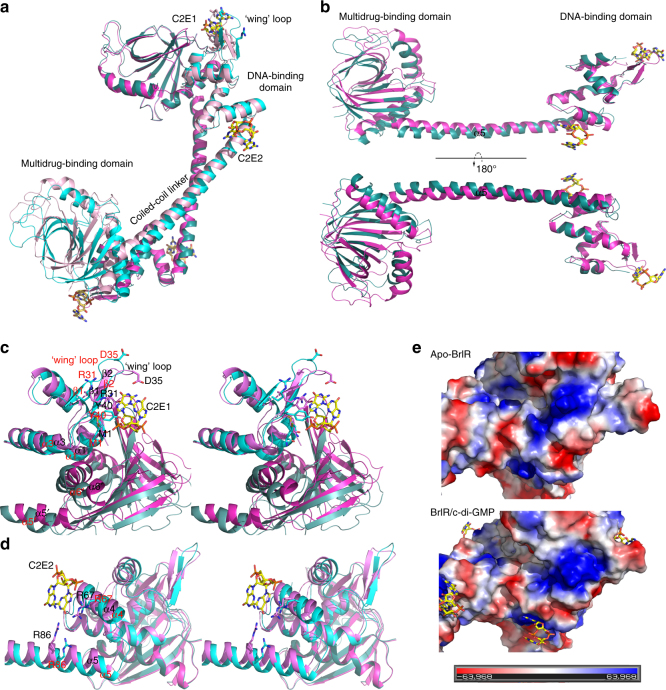
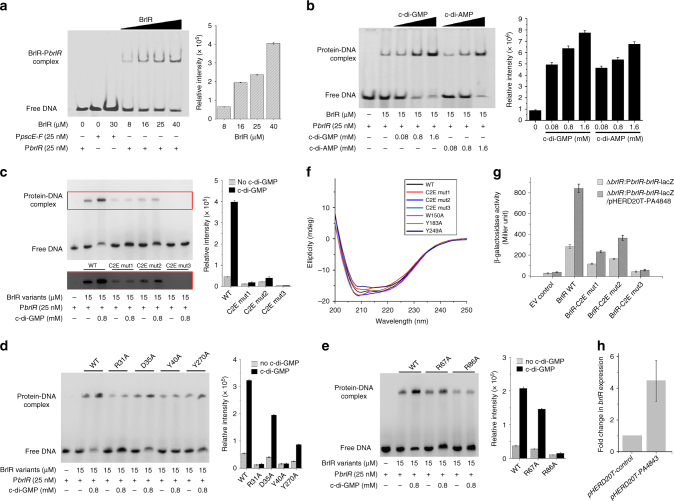
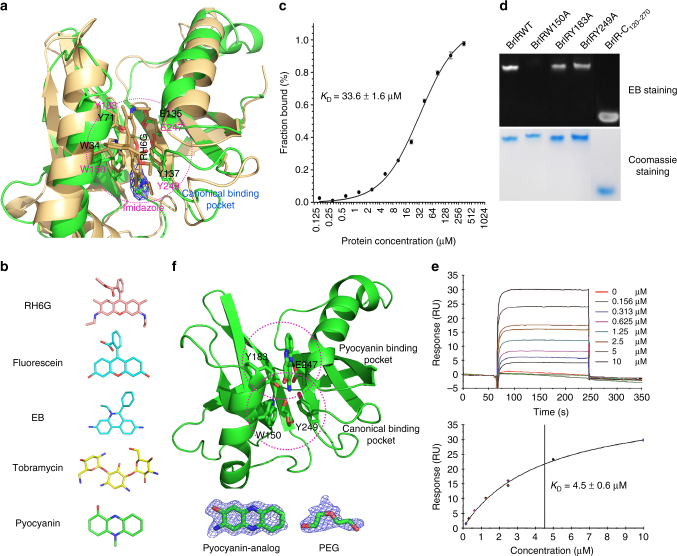
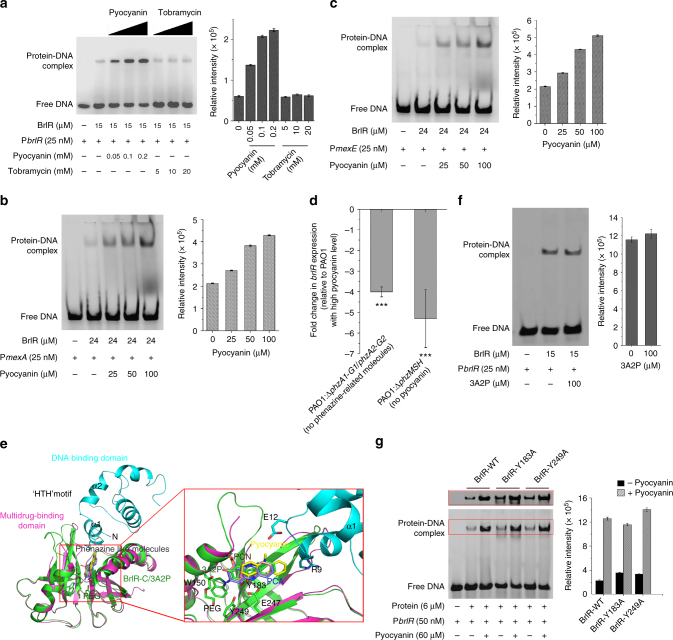
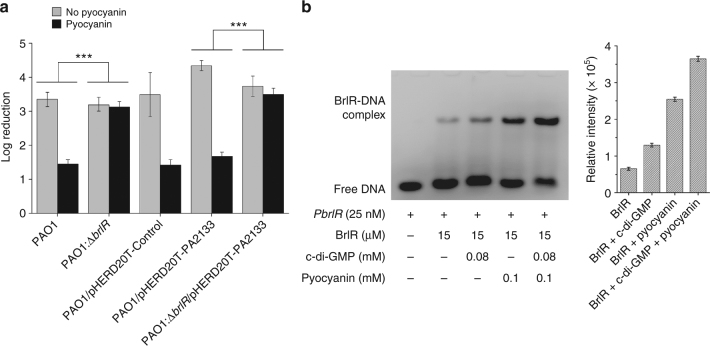
The virulence factor pyocyanin and the intracellular second messenger cyclic diguanylate monophosphate (c-di-GMP) play key roles in regulating biofilm formation and multi-drug efflux pump expression in Pseudomonas aeruginosa. However, the crosstalk between these two signaling pathways remains unclear. Here we show that BrlR (PA4878), previously identified as a c-di-GMP responsive transcriptional regulator, acts also as a receptor for pyocyanin. Crystal structures of free BrlR and c-di-GMP-bound BrlR reveal that the DNA-binding domain of BrlR contains two separate c-di-GMP binding sites, both of which are involved in promoting brlR expression. In addition, we identify a pyocyanin-binding site on the C-terminal multidrug-binding domain based on the structure of the BrlR-C domain in complex with a pyocyanin analog. Biochemical analysis indicates that pyocyanin enhances BrlR-DNA binding and brlR expression in a concentration-dependent manner.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
