

Structure of the core of the type III secretion system export apparatus
- PMID: 29967543
- PMCID: PMC6233869
- DOI: 10.1038/s41594-018-0086-9
Structure of the core of the type III secretion system export apparatus
Erratum in
-
Author Correction: Structure of the core of the type III secretion system export apparatus.Nat Struct Mol Biol. 2018 Aug;25(8):743. doi: 10.1038/s41594-018-0095-8. Nat Struct Mol Biol. 2018. PMID: 30018321
Abstract
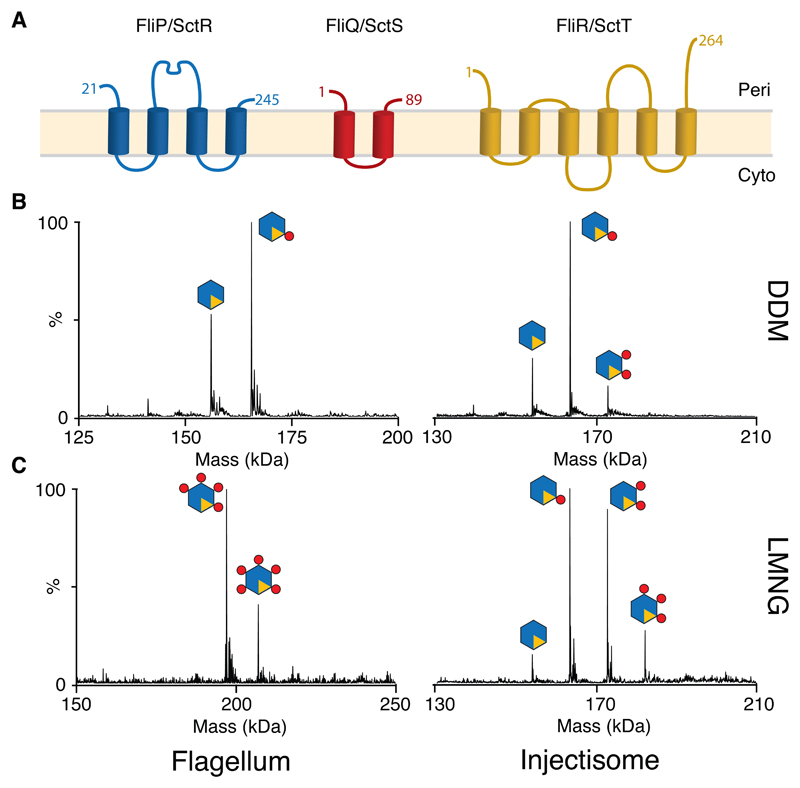
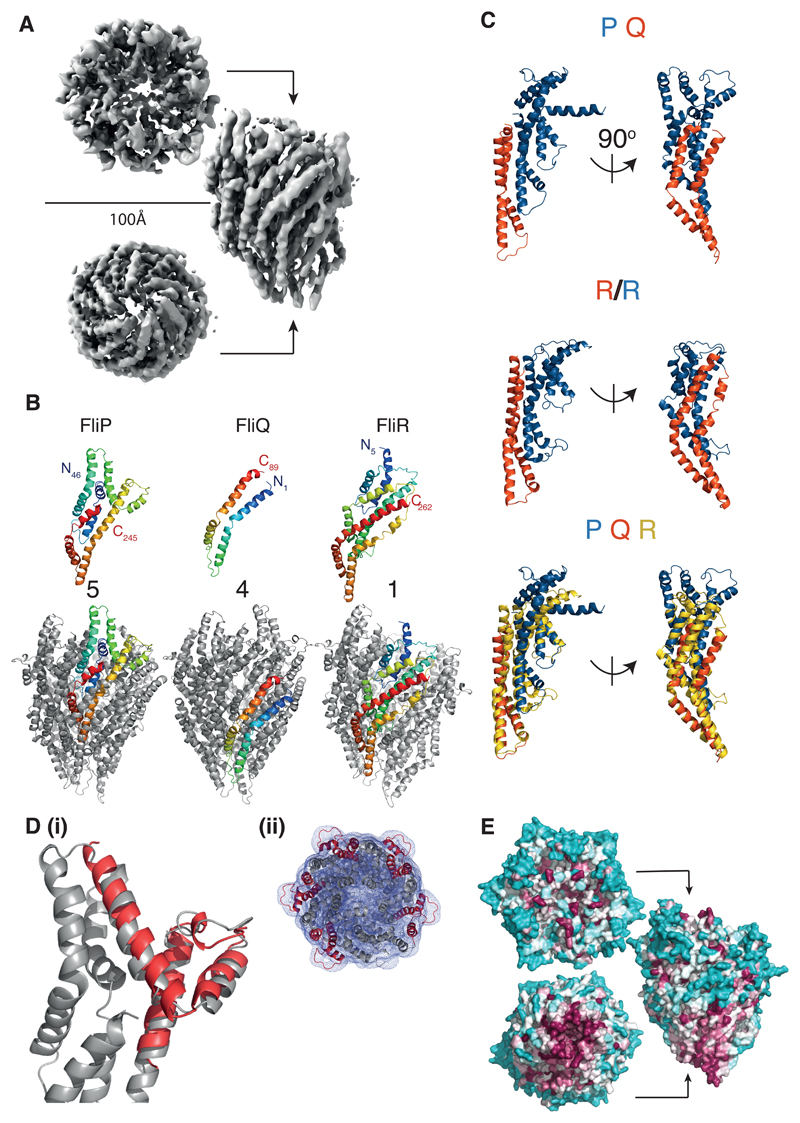
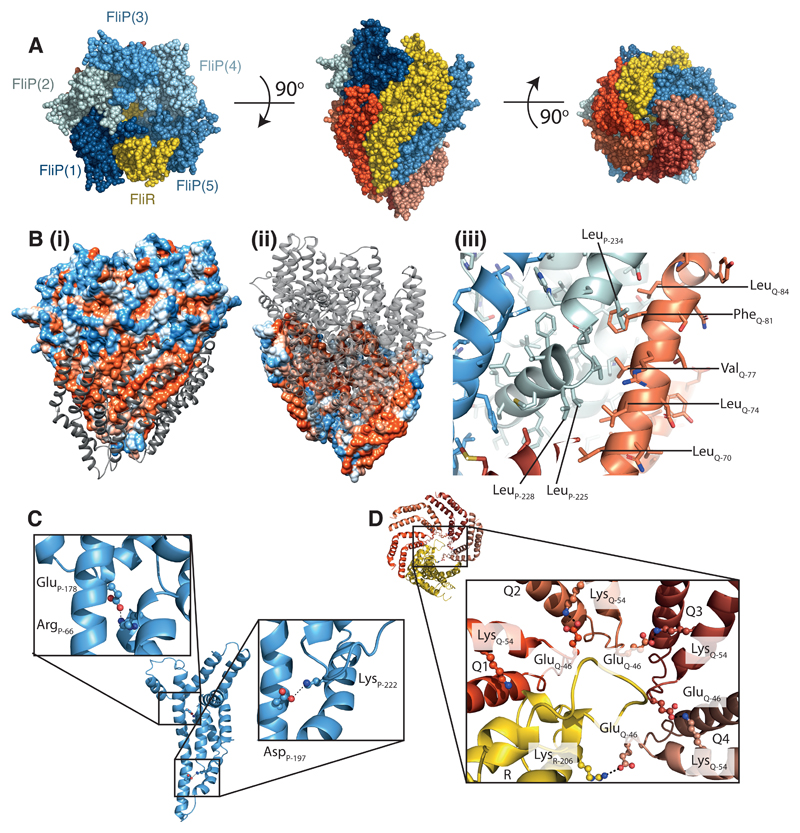
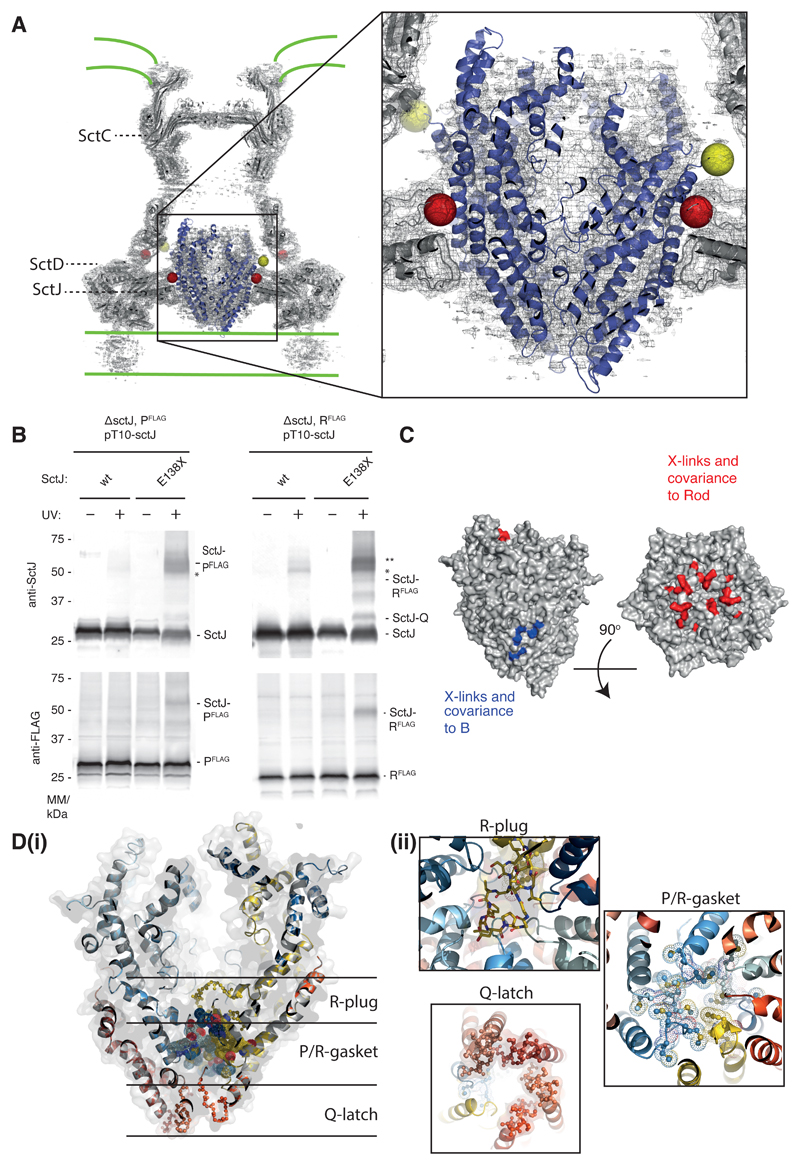
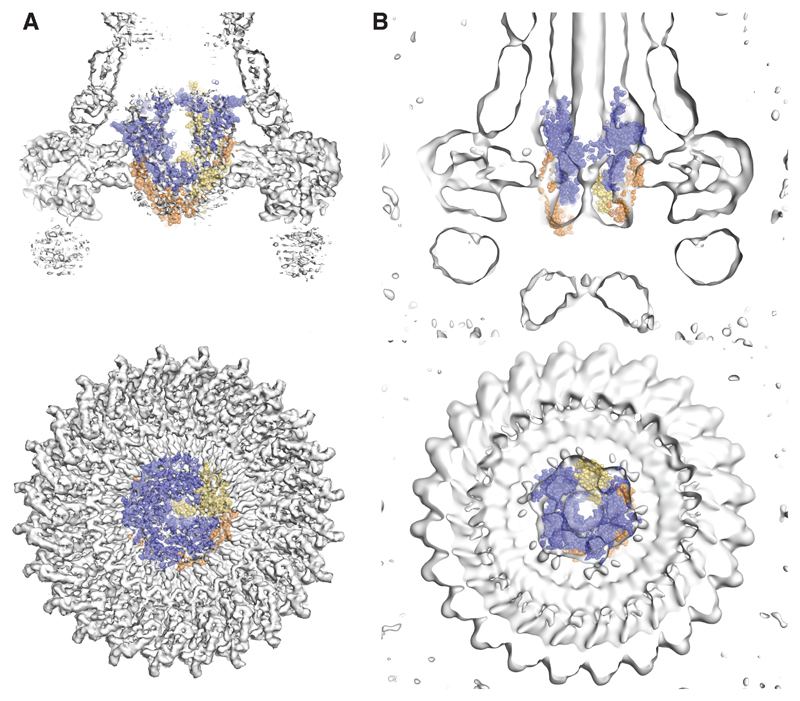
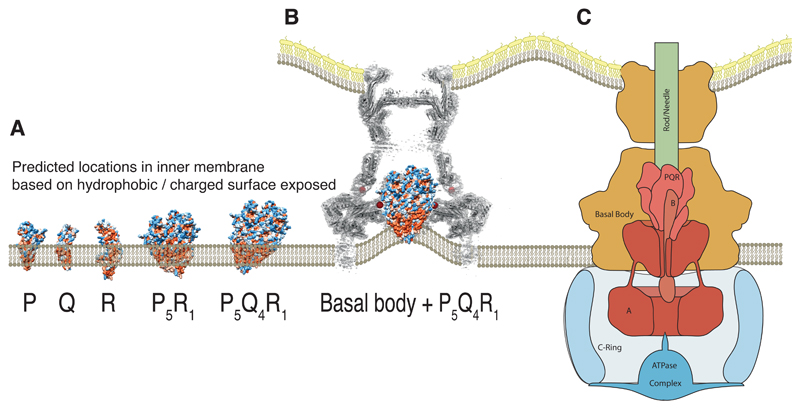
Export of proteins through type III secretion systems is critical for motility and virulence of many major bacterial pathogens. Three putative integral membrane proteins (FliP, FliQ, FliR) are suggested to form the core of an export gate in the inner membrane, but their structure, assembly and location within the final nanomachine remain unclear. Here, we present the cryoelectron microscopy structure of the Salmonella Typhimurium FliP-FliQ-FliR complex at 4.2 Å. None of the subunits adopt canonical integral membrane protein topologies, and common helix-turn-helix structural elements allow them to form a helical assembly with 5:4:1 stoichiometry. Fitting of the structure into reconstructions of intact secretion systems, combined with cross-linking, localize the export gate as a core component of the periplasmic portion of the machinery. This study thereby identifies the export gate as a key element of the secretion channel and implies that it primes the helical architecture of the components assembling downstream.
Figures
Similar articles
-
High-resolution view of the type III secretion export apparatus in situ reveals membrane remodeling and a secretion pathway.Proc Natl Acad Sci U S A. 2019 Dec 3;116(49):24786-24795. doi: 10.1073/pnas.1916331116. Epub 2019 Nov 19. Proc Natl Acad Sci U S A. 2019. PMID: 31744874 Free PMC article.
-
The Structure of an Injectisome Export Gate Demonstrates Conservation of Architecture in the Core Export Gate between Flagellar and Virulence Type III Secretion Systems.mBio. 2019 Jun 25;10(3):e00818-19. doi: 10.1128/mBio.00818-19. mBio. 2019. PMID: 31239376 Free PMC article.
-
Determination of the Stoichiometry of the Complete Bacterial Type III Secretion Needle Complex Using a Combined Quantitative Proteomic Approach.Mol Cell Proteomics. 2016 May;15(5):1598-609. doi: 10.1074/mcp.M115.056598. Epub 2016 Feb 21. Mol Cell Proteomics. 2016. PMID: 26900162 Free PMC article.
-
Structures of Type III Secretion System Needle Filaments.Curr Top Microbiol Immunol. 2020;427:109-131. doi: 10.1007/82_2019_192. Curr Top Microbiol Immunol. 2020. PMID: 31974760 Review.
-
Molecular Organization and Assembly of the Export Apparatus of Flagellar Type III Secretion Systems.Curr Top Microbiol Immunol. 2020;427:91-107. doi: 10.1007/82_2019_170. Curr Top Microbiol Immunol. 2020. PMID: 31172377 Review.
Cited by
-
A positive charge region of Salmonella FliI is required for ATPase formation and efficient flagellar protein export.Commun Biol. 2021 Apr 12;4(1):464. doi: 10.1038/s42003-021-01980-y. Commun Biol. 2021. PMID: 33846530 Free PMC article.
-
Substrate-engaged type III secretion system structures reveal gating mechanism for unfolded protein translocation.Nat Commun. 2021 Mar 9;12(1):1546. doi: 10.1038/s41467-021-21143-1. Nat Commun. 2021. PMID: 33750771 Free PMC article.
-
The Missing Pieces: The Role of Secretion Systems in Campylobacter jejuni Virulence.Biomolecules. 2023 Jan 9;13(1):135. doi: 10.3390/biom13010135. Biomolecules. 2023. PMID: 36671522 Free PMC article. Review.
-
Control of membrane barrier during bacterial type-III protein secretion.Nat Commun. 2021 Jun 28;12(1):3999. doi: 10.1038/s41467-021-24226-1. Nat Commun. 2021. PMID: 34183670 Free PMC article.
-
Establishment of Nested PCR for the Detection of Pseudomonas plecoglossicida and Epidemiological Survey of Larimichthys crocea in the Southeast Coastal Region.Animals (Basel). 2024 May 10;14(10):1427. doi: 10.3390/ani14101427. Animals (Basel). 2024. PMID: 38791645 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
