

Peptides derived from the integrin β cytoplasmic tails inhibit angiogenesis
- PMID: 29970081
- PMCID: PMC6029062
- DOI: 10.1186/s12964-018-0248-8
Peptides derived from the integrin β cytoplasmic tails inhibit angiogenesis
Abstract
Background: Integrins are essential regulators of angiogenesis. However, the antiangiogenic potential of peptides derived from the integrin cytoplasmic tails (CT) remains mostly undetermined.
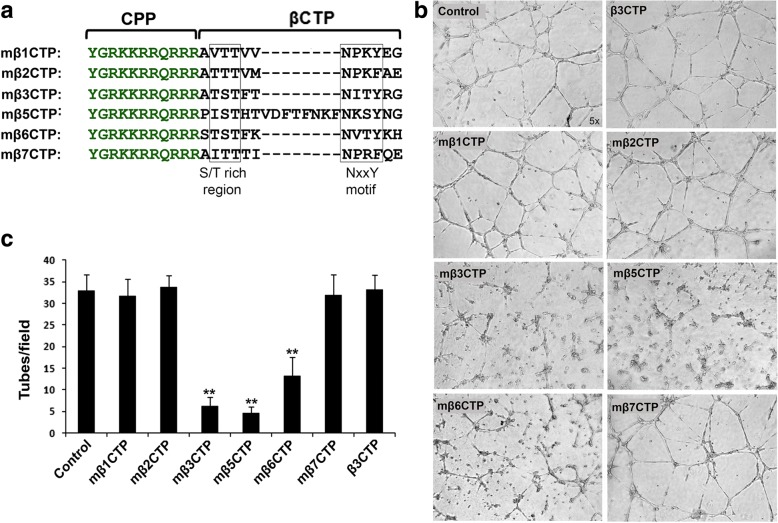
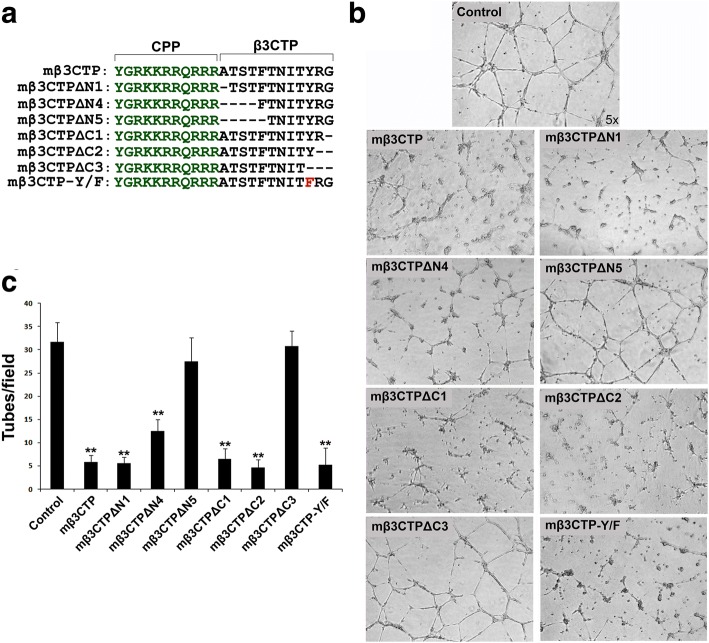
Methods: Here we designed a panel of membrane-penetrating peptides (termed as mβCTPs), each comprising a C-terminal NxxY motif from one of the conserved integrin β CTs, and evaluated their antiangiogenic ability using both in vitro and in vivo approaches.
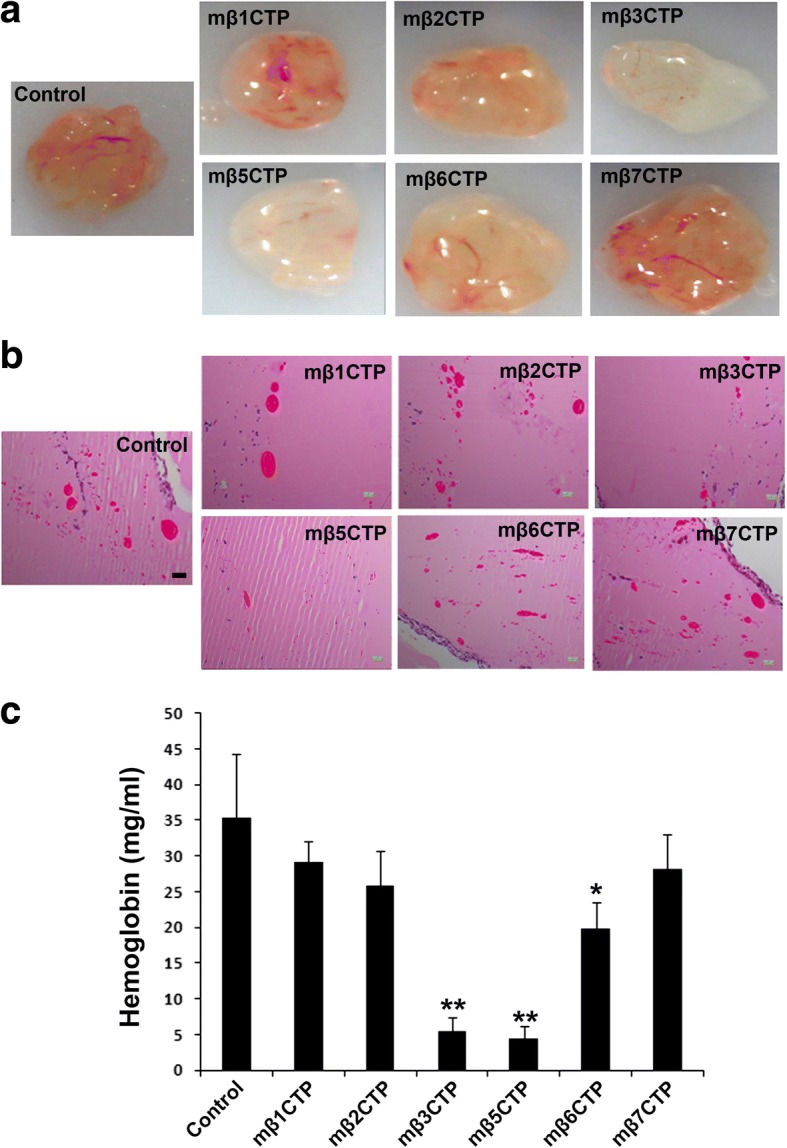
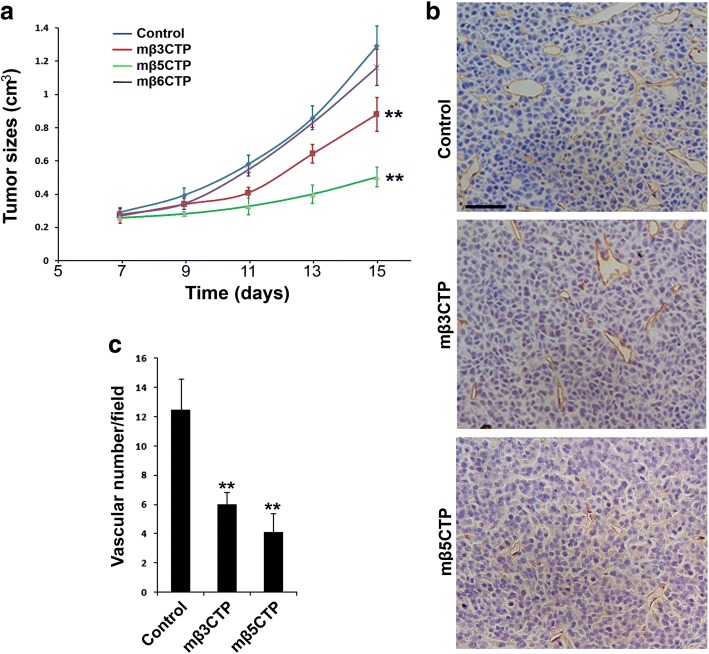
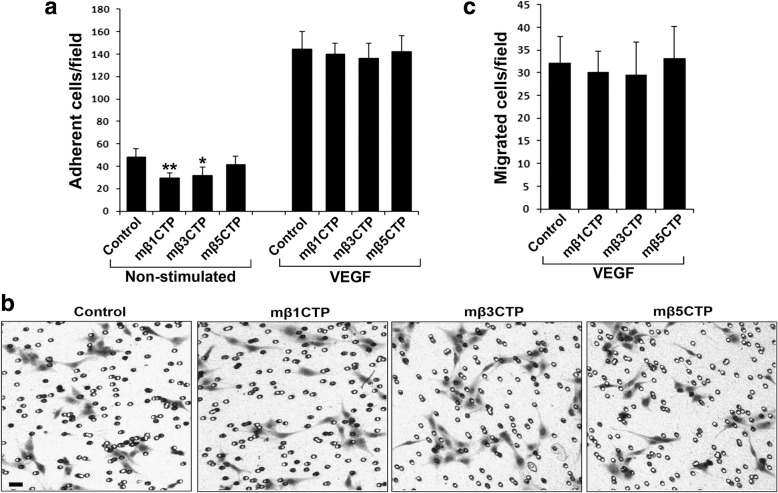
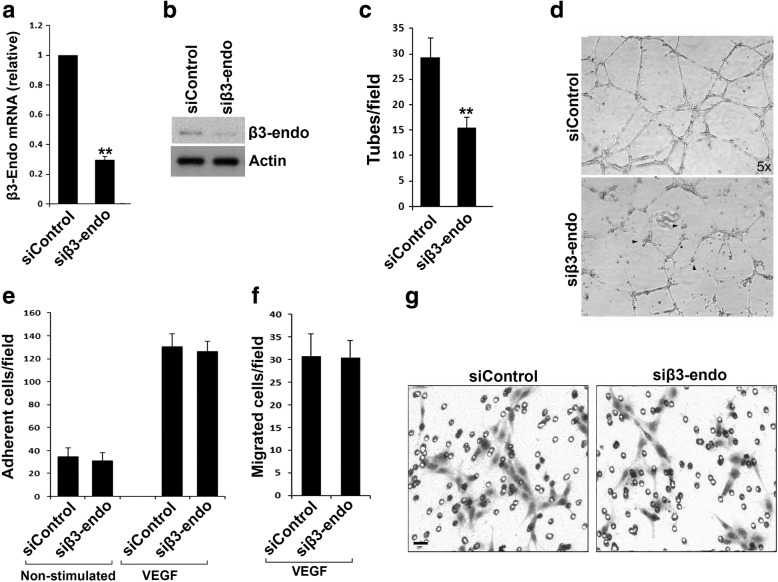
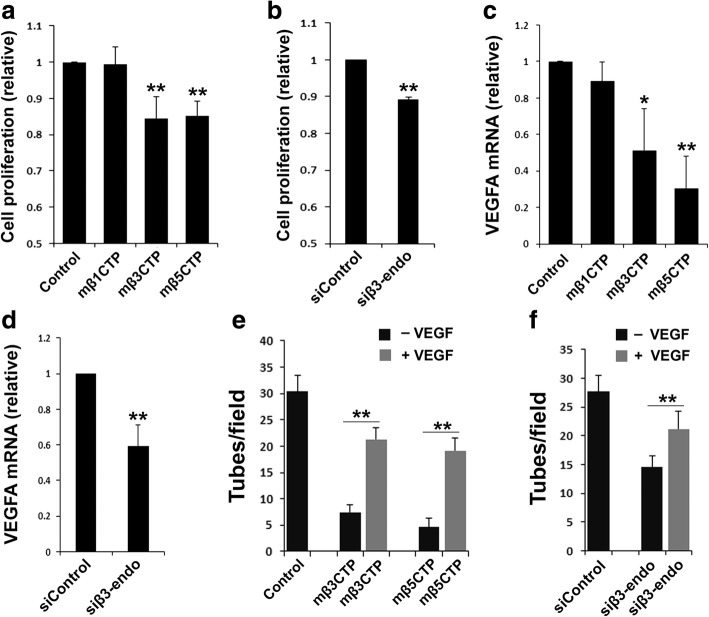
Results: We found that mβ3CTP, mβ5CTP and mβ6CTP, derived respectively from the integrin β3, β5 and β6 CTs, but not others, exhibit antiangiogenic ability. Interestingly, we observed that the integrin β3, β5 and β6 CTs but not others are able to interact with β3-endonexin. In addition, the antiangiogenic core in mβ3CTP is identical to a previously identified β3-endonexin binding region in the integrin β3 CT, indicating that the antiangiogenic mβCTPs may function via their binding to β3-endonexin. Consistently, knockdown of endogenous β3-endonexin in HUVECs significantly suppresses tube formation, suggesting that β3-endonexin is proangiogenic. However, neither treatment with the antiangiogenic mβCTPs nor knockdown of endogenous β3-endonexin affects integrin-mediated HUVEC adhesion and migration, indicating that their antiangiogenic effect may not rely on directly regulating integrin activity. Importantly, both treatment with the antiangiogenic mβCTPs and knockdown of endogenous β3-endonexin in HUVECs inhibit VEGF expression and cell proliferation, thereby providing mechanistic explanations for the functional consequences.
Conclusion: Our results suggest that the antiangiogenic mβCTPs can interact with β3-endonexin in vascular endothelial cells and suppress its function in regulating VEGF expression and cell proliferation, thus disclosing a unique pathway that may be useful for developing novel antiangiogenic strategies.
Keywords: Angiogenesis; Cytoplasmic tails; Endothelial cells; Integrins; β3-endonexin.
Conflict of interest statement
Ethics approval and consent to participate
All animal handling and experiments were approved by the IACUC of Shanghai University School of Life Sciences.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures

Similar articles
-
β3-Endonexin interacts with ninein in vascular endothelial cells to promote angiogenesis.Biochem Biophys Res Commun. 2021 Aug 20;566:75-79. doi: 10.1016/j.bbrc.2021.06.008. Epub 2021 Jun 9. Biochem Biophys Res Commun. 2021. PMID: 34118594
-
A conserved sequence motif in the integrin beta3 cytoplasmic domain is required for its specific interaction with beta3-endonexin.J Biol Chem. 1997 Mar 21;272(12):7693-8. doi: 10.1074/jbc.272.12.7693. J Biol Chem. 1997. PMID: 9065427
-
Interaction Analyses of 14-3-3ζ, Dok1, and Phosphorylated Integrin β Cytoplasmic Tails Reveal a Bi-molecular Switch in Integrin Regulation.J Mol Biol. 2018 Oct 19;430(21):4419-4430. doi: 10.1016/j.jmb.2018.09.008. Epub 2018 Sep 20. J Mol Biol. 2018. PMID: 30243836
-
Integrin cytoplasmic interactions and bidirectional transmembrane signalling.Curr Opin Cell Biol. 1996 Oct;8(5):657-69. doi: 10.1016/s0955-0674(96)80107-4. Curr Opin Cell Biol. 1996. PMID: 8939656 Review.
-
Conserved functions of the cytoplasmic domains of integrin beta subunits.Front Biosci. 1998 Aug 5;3:d877-86. doi: 10.2741/a329. Front Biosci. 1998. PMID: 9693157 Review.
Cited by
-
High Density Display of an Anti-Angiogenic Peptide on Micelle Surfaces Enhances Their Inhibition of αvβ3 Integrin-Mediated Neovascularization In Vitro.Nanomaterials (Basel). 2020 Mar 22;10(3):581. doi: 10.3390/nano10030581. Nanomaterials (Basel). 2020. PMID: 32235802 Free PMC article.
-
Paxillin binding to the PH domain of kindlin-3 in platelets is required to support integrin αIIbβ3 outside-in signaling.J Thromb Haemost. 2021 Dec;19(12):3126-3138. doi: 10.1111/jth.15505. Epub 2021 Aug 31. J Thromb Haemost. 2021. PMID: 34411430 Free PMC article.
-
In vitro assessment of varying peptide surface density on the suppression of angiogenesis by micelles displaying αvβ3 blocking peptides.J Biomed Mater Res B Appl Biomater. 2023 Feb;111(2):343-353. doi: 10.1002/jbm.b.35154. Epub 2022 Aug 30. J Biomed Mater Res B Appl Biomater. 2023. PMID: 36054456 Free PMC article.
-
The MemMoRF database for recognizing disordered protein regions interacting with cellular membranes.Nucleic Acids Res. 2021 Jan 8;49(D1):D355-D360. doi: 10.1093/nar/gkaa954. Nucleic Acids Res. 2021. PMID: 33119751 Free PMC article.
-
Understanding the role of integrins in breast cancer invasion, metastasis, angiogenesis, and drug resistance.Oncogene. 2021 Feb;40(6):1043-1063. doi: 10.1038/s41388-020-01588-2. Epub 2021 Jan 8. Oncogene. 2021. PMID: 33420366 Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
