

PATs and SNATs: Amino Acid Sensors in Disguise
- PMID: 29971004
- PMCID: PMC6018406
- DOI: 10.3389/fphar.2018.00640
PATs and SNATs: Amino Acid Sensors in Disguise
Abstract
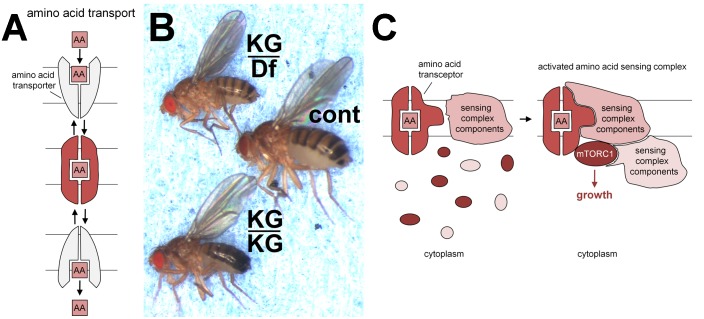
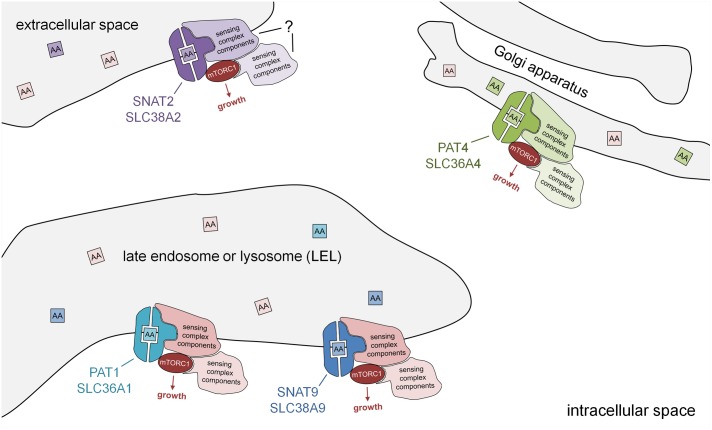
Solute Carriers (SLCs) are involved in the transport of substances across lipid bilayers, including nutrients like amino acids. Amino acids increase the activity of the microenvironmental sensor mechanistic Target of Rapamycin Complex 1 (mTORC1) to promote cellular growth and anabolic processes. They can be brought in to cells by a wide range of SLCs including the closely related Proton-assisted Amino acid Transporter (PAT or SLC36) and Sodium-coupled Neutral Amino acid Transporter (SNAT or SLC38) families. More than a decade ago, the first evidence emerged that members of the PAT family can act as amino acid-stimulated receptors, or so-called "transceptors," connecting amino acids to mTORC1 activation. Since then, further studies in human cell models have suggested that other PAT and SNAT family members, which share significant homology within their transmembrane domains, can act as transceptors. A paradigm shift has also led to the PATs and SNATs at the surface of multiple intracellular compartments being linked to the recruitment and activation of different pools of mTORC1. Much focus has been on late endosomes and lysosomes as mTORC1 regulatory hubs, but more recently a Golgi-localized PAT was shown to be required for mTORC1 activation. PATs and SNATs can also traffic between the cell surface and intracellular compartments, with regulation of this movement providing a means of controlling their mTORC1 regulatory activity. These emerging features of PAT and SNAT amino acid sensors, including the transceptor mechanism, have implications for the pharmacological inhibition of mTORC1 and new therapeutic interventions.
Keywords: SLC36A1; SLC36A4; SLC38A9; SNAT2; amino acid transporter; mechanistic target of rapamycin (mTORC1); transceptor.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases