

Dipeptide repeat proteins activate a heat shock response found in C9ORF72-ALS/FTLD patients
- PMID: 29973287
- PMCID: PMC6031111
- DOI: 10.1186/s40478-018-0555-8
Dipeptide repeat proteins activate a heat shock response found in C9ORF72-ALS/FTLD patients
Abstract
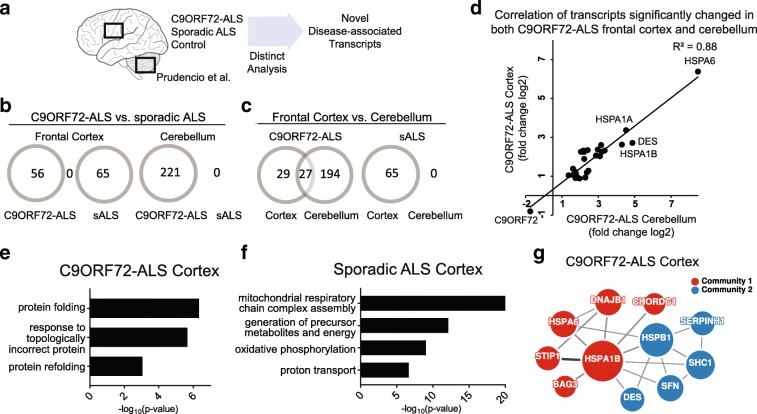
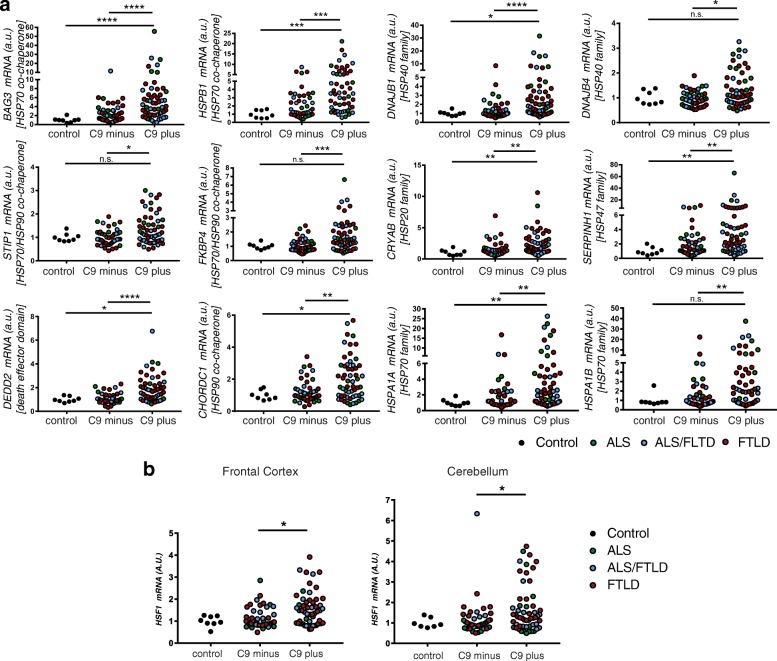
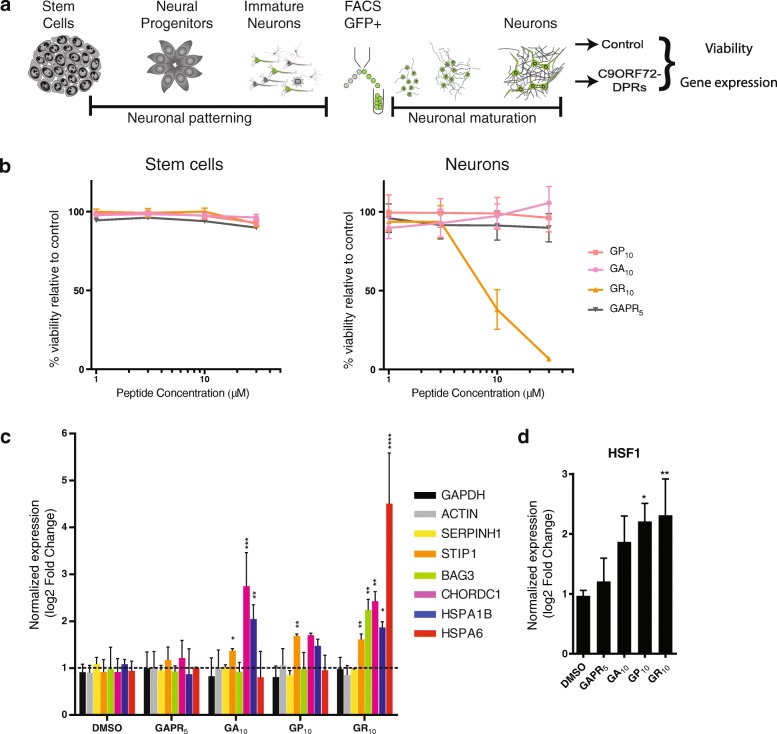
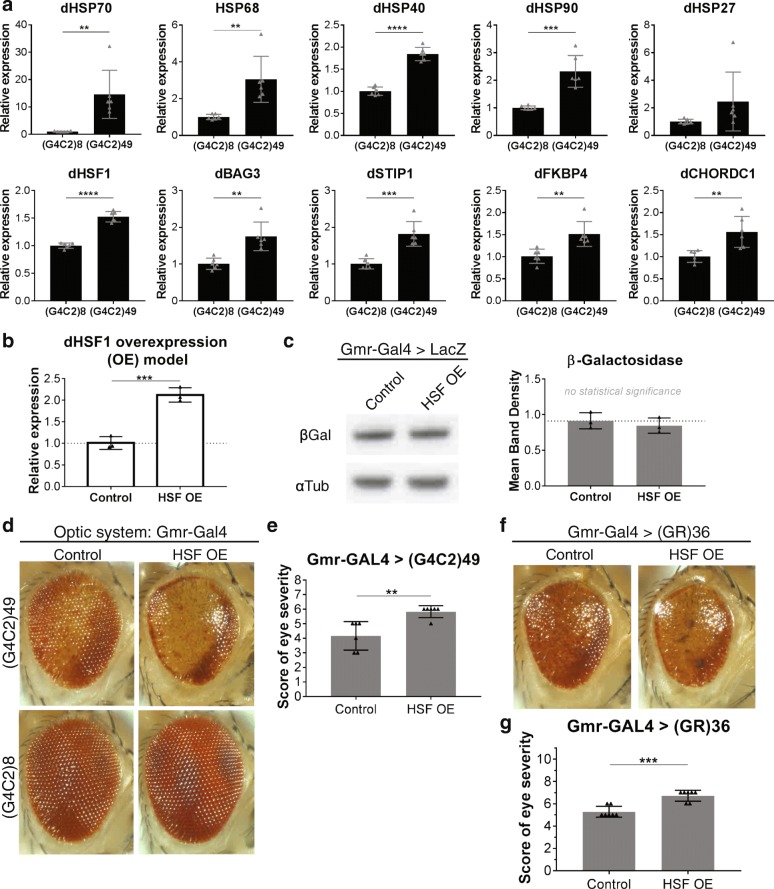
A hexanucleotide (GGGGCC) repeat expansion in C9ORF72 is the most common genetic contributor to amyotrophic lateral sclerosis (ALS) and frontotemporal lobar degeneration (FTLD). Reduced expression of the C9ORF72 gene product has been proposed as a potential contributor to disease pathogenesis. Additionally, repetitive RNAs and dipeptide repeat proteins (DPRs), such as poly-GR, can be produced by this hexanucleotide expansion that disrupt a number of cellular processes, potentially contributing to neural degeneration. To better discern which of these mechanisms leads to disease-associated changes in patient brains, we analyzed gene expression data generated from the cortex and cerebellum. We found that transcripts encoding heat shock proteins (HSPs) regulated by the HSF1 transcription factor were significantly induced in C9ORF72-ALS/FTLD patients relative to both sporadic ALS/FTLD cases and controls. Treatment of human neurons with chemically synthesized DPRs was sufficient to activate a similar transcriptional response. Expression of GGGGCC repeats and also poly-GR in the brains of Drosophila lead to the upregulation of HSF1 and the same highly-conserved HSPs. Additionally, HSF1 was a modifier of poly-GR toxicity in Drosophila. Our results suggest that the expression of DPRs are associated with upregulation of HSF1 and activation of a heat shock response in C9ORF72-ALS/FTLD.
Keywords: Amyotrophic lateral sclerosis; C9ORF72 repeat expansion; Dipeptide repeat proteins; Drosophila; Frontotemporal dementia; Frontotemporal lobar degeneration; HSF1; Heat shock response.
Conflict of interest statement
Ethics approval and consent to participate
Written informed consent was given by all participants or authorized family members in accordance with institutional review board guidelines of Mayo Clinic.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Agosta F, Ferraro PM, Riva N, Spinelli EG, Domi T, Carrera P, Copetti M, Falzone Y, Ferrari M, Lunetta C, Comi G, Falini A, Quattrini A, Filippi M (2017) Structural and functional brain signatures of C9orf72 in motor neuron disease. Neurobiol Aging. 10.1016/j.neurobiolaging.2017.05.024 - PubMed
-
- Al-Sarraj S, King A, Troakes C, Smith B, Maekawa S, Bodi I, Rogelj B, Al-Chalabi A, Hortobagyi T, Shaw CE. p62 positive, TDP-43 negative, neuronal cytoplasmic and intranuclear inclusions in the cerebellum and hippocampus define the pathology of C9orf72-linked FTLD and MND/ALS. Acta Neuropathol. 2011;122:691–702. doi: 10.1007/s00401-011-0911-2. - DOI - PubMed
-
- Berson A, Sartoris A, Nativio R, Van Deerlin V, Toledo JB, Porta S, Liu S, Chung CY, Garcia BA, Lee VM, Trojanowski JQ, Johnson FB, Berger SL, Bonini NM (2017) TDP-43 promotes neurodegeneration by impairing chromatin remodeling. Curr Biol 27:3579–3590 e3576. doi:10.1016/j.cub.2017.10.024 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS089742/NS/NINDS NIH HHS/United States
- R35 NS097273/NS/NINDS NIH HHS/United States
- P50 AG016574/AG/NIA NIH HHS/United States
- P30 AG062421/AG/NIA NIH HHS/United States
- P01 NS084974/NS/NINDS NIH HHS/United States
- R21 NS084528/NS/NINDS NIH HHS/United States
- R01 ES020395/ES/NIEHS NIH HHS/United States
- R35 NS097275/NS/NINDS NIH HHS/United States
- R01 NS077402/NS/NINDS NIH HHS/United States
- R01 NS063964/NS/NINDS NIH HHS/United States
- P50 AG005134/AG/NIA NIH HHS/United States
- T32 CA009216/CA/NCI NIH HHS/United States
- R01 NS088689/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
