

microRNA-122 amplifies hepatitis C virus translation by shaping the structure of the internal ribosomal entry site
- PMID: 29973597
- PMCID: PMC6031695
- DOI: 10.1038/s41467-018-05053-3
microRNA-122 amplifies hepatitis C virus translation by shaping the structure of the internal ribosomal entry site
Abstract
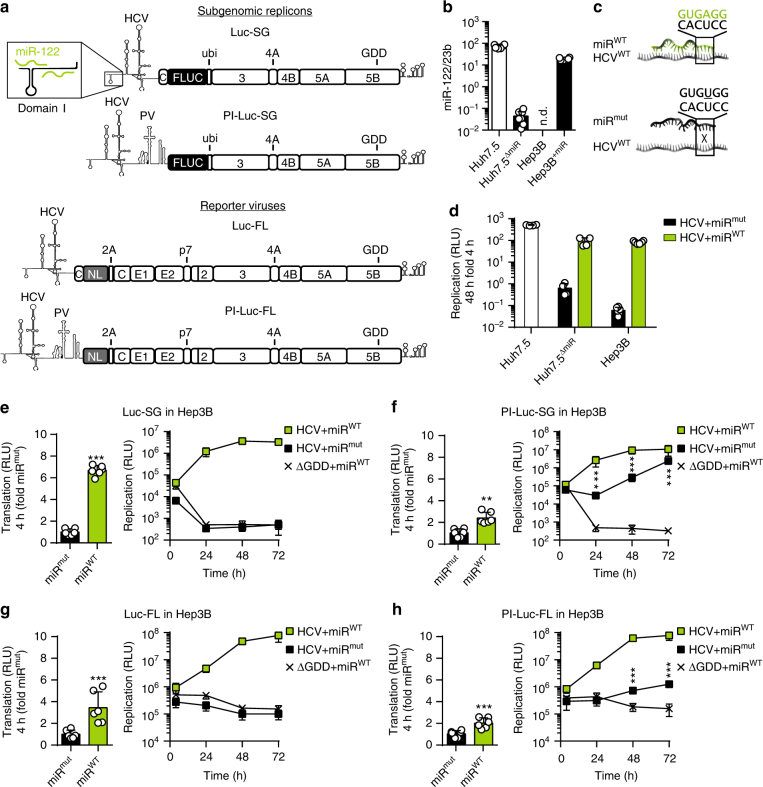
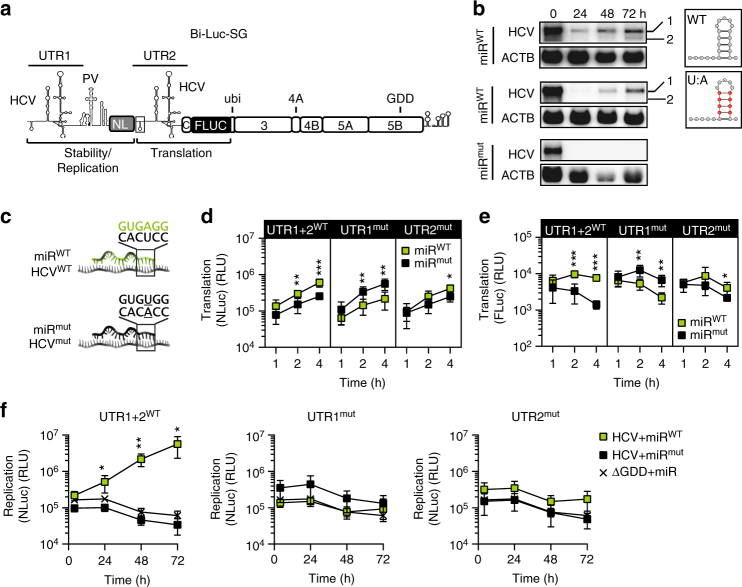
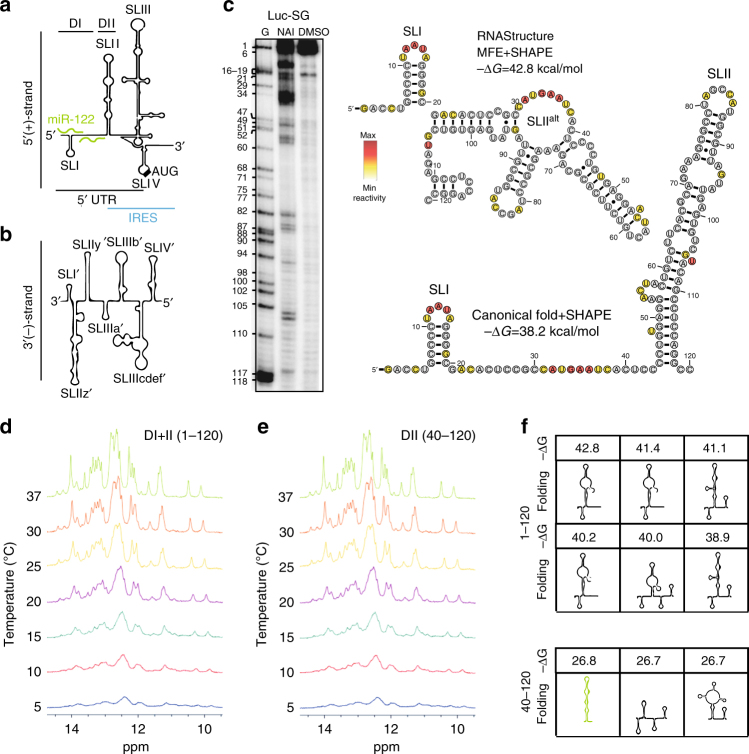
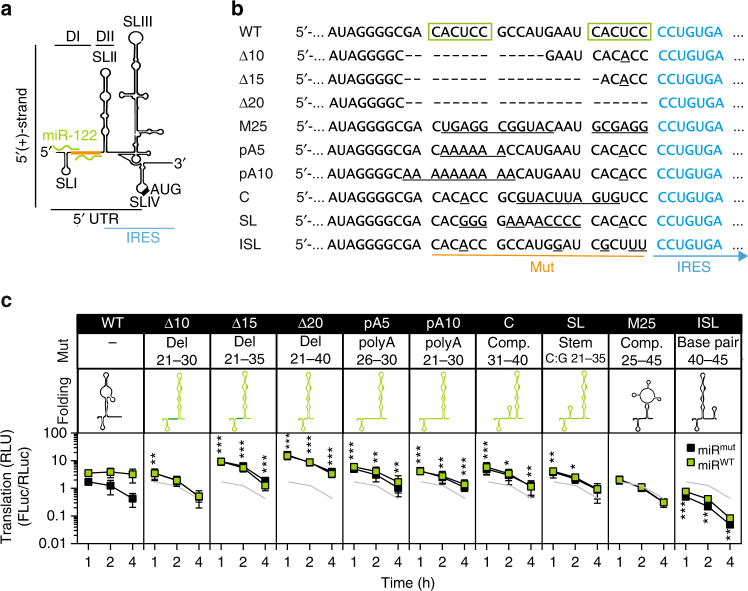
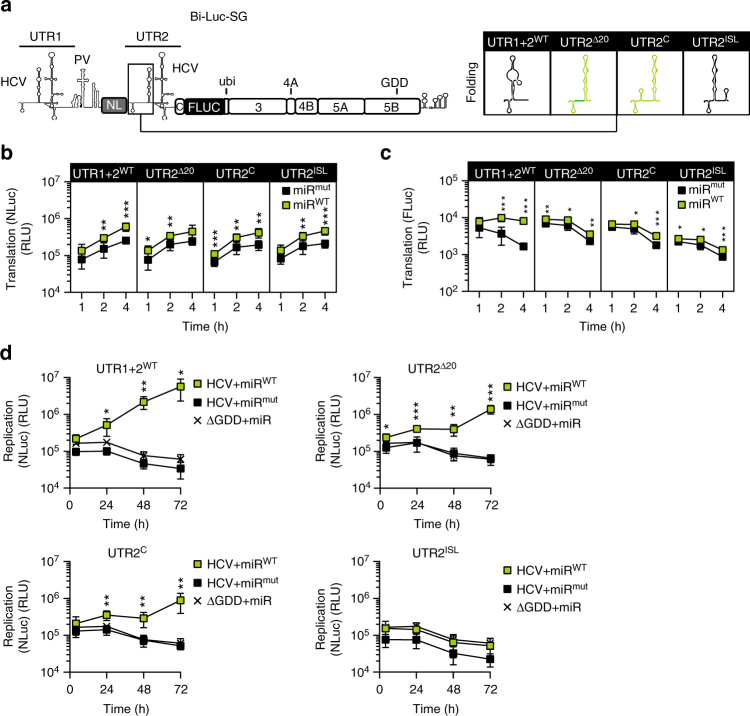
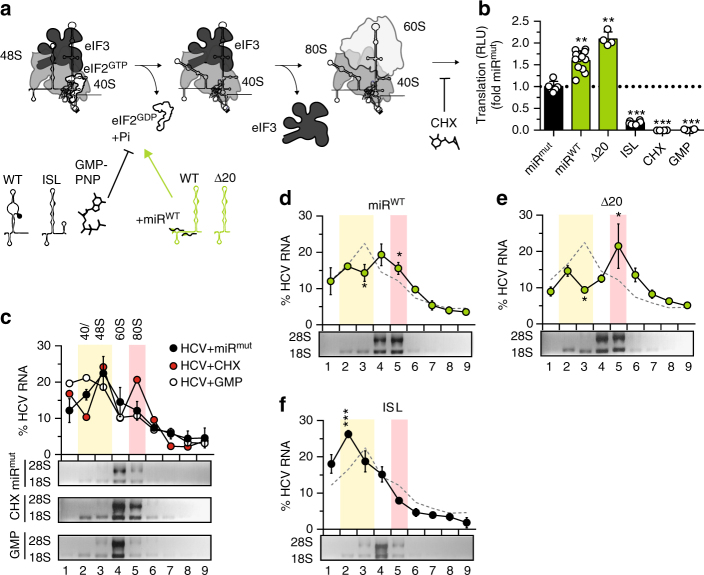
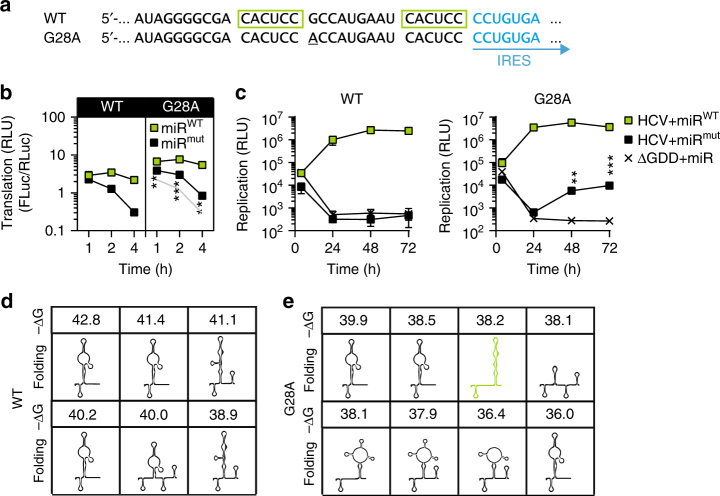
The liver-specific microRNA-122 (miR-122) recognizes two conserved sites at the 5' end of the hepatitis C virus (HCV) genome and contributes to stability, translation, and replication of the viral RNA. We show that stimulation of the HCV internal ribosome entry site (IRES) by miR-122 is essential for efficient viral replication. The mechanism relies on a dual function of the 5' terminal sequence in the complementary positive (translation) and negative strand (replication), requiring different secondary structures. Predictions and experimental evidence argue for several alternative folds involving the miR-binding region (MBR) adjacent to the IRES and interfering with its function. Mutations in the MBR, designed to suppress these dysfunctional structures indeed stimulate translation independently of miR-122. Conversely, MBR mutants favoring alternative folds show impaired IRES activity. Our results therefore suggest that miR-122 binding assists the folding of a functional IRES in an RNA chaperone-like manner by suppressing energetically favorable alternative secondary structures.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
