

An Engineered Distant Homolog of Pseudomonas syringae TTSS Effector From Physcomitrella patens Can Act as a Bacterial Virulence Factor
- PMID: 29973916
- PMCID: PMC6019455
- DOI: 10.3389/fmicb.2018.01060
An Engineered Distant Homolog of Pseudomonas syringae TTSS Effector From Physcomitrella patens Can Act as a Bacterial Virulence Factor
Abstract
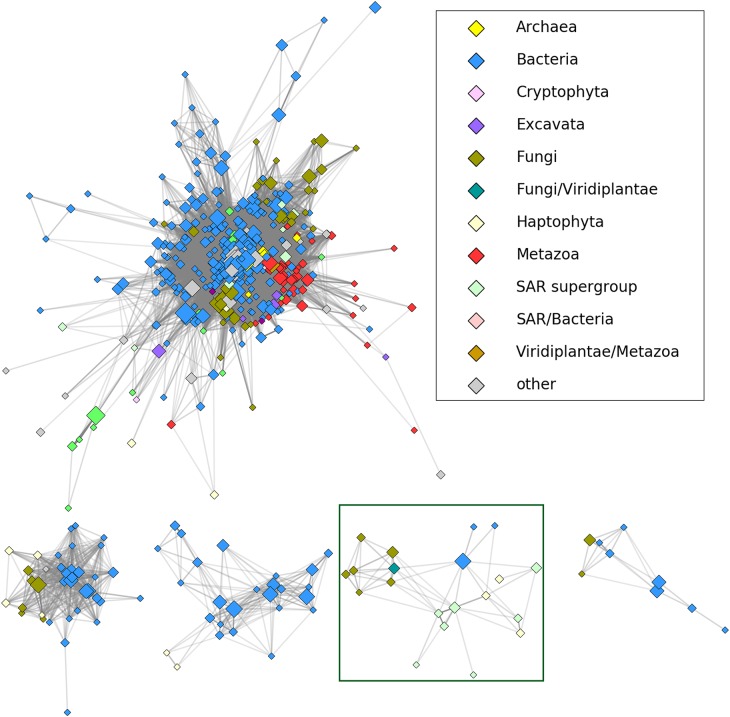
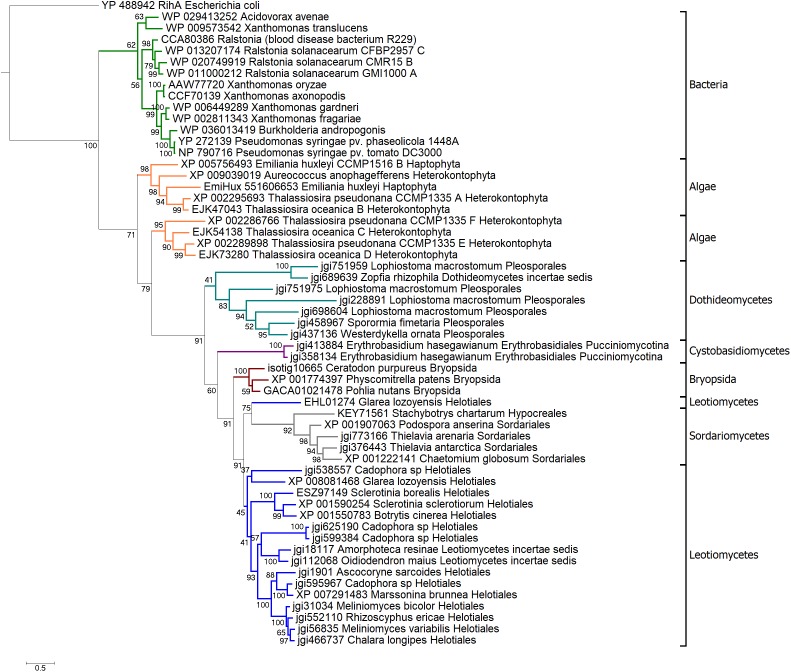
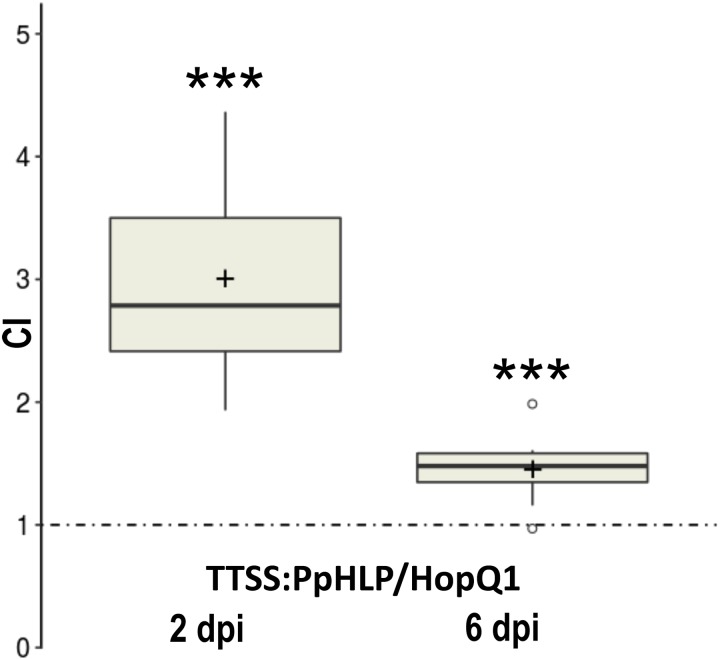
Pseudomonas syringae pv. phaseolicola is the causative agent of halo blight in common bean (Phaseolus vulgaris). Similar to other pathogenic gram-negative bacteria, it secrets a set of type III effectors into host cells to subvert defense mechanisms. HopQ1 (for Hrp outer protein Q) is one of these type III effectors contributing to virulence of bacteria. Upon delivery into a plant cell, HopQ1 undergoes phosphorylation, binds host 14-3-3 proteins and suppresses defense-related signaling. Some plants however, evolved systems to recognize HopQ1 and respond to its presence and thus to prevent infection. HopQ1 shows homology to Nucleoside Hydrolases (NHs), but it contains a modified calcium binding motif not found in the canonical enzymes. CLuster ANalysis of Sequences (CLANS) revealed that HopQ1 and alike proteins make a distinct group of putative NHs located distantly from the classical enzymes. The HopQ1 - like protein (HLP) group comprises sequences from plant pathogenic bacteria, fungi, and lower plants. Our data suggest that the evolution of HopQ1 homologs in bacteria, fungi, and algae was independent. The location of moss HopQ1 homologs inside the fungal clade indicates a possibility of horizontal gene transfer (HGT) between those taxa. We identified a HLP in the moss Physcomitrella patens. Our experiments show that this protein (referred to as PpHLP) extended by a TTSS signal of HopQ1 promoted P. syringae growth in bean and was recognized by Nicotiana benthamiana immune system. Thus, despite the low sequence similarity to HopQ1 the engineered PpHLP acted as a bacterial virulence factor and displayed similar to HopQ1 virulence properties.
Keywords: HopQ1; Physcomitrella patens; Pseudomonas syringae; evolution; horizontal gene transfer; type three effector.
Figures

Similar articles
-
Phosphorylation of HopQ1, a type III effector from Pseudomonas syringae, creates a binding site for host 14-3-3 proteins.Plant Physiol. 2013 Apr;161(4):2049-61. doi: 10.1104/pp.112.209023. Epub 2013 Feb 8. Plant Physiol. 2013. PMID: 23396834 Free PMC article.
-
Two Strategies of Pseudomonas syringae to Avoid Recognition of the HopQ1 Effector in Nicotiana Species.Front Plant Sci. 2018 Jul 10;9:978. doi: 10.3389/fpls.2018.00978. eCollection 2018. Front Plant Sci. 2018. PMID: 30042777 Free PMC article.
-
Contributions of the effector gene hopQ1-1 to differences in host range between Pseudomonas syringae pv. phaseolicola and P. syringae pv. tabaci.Mol Plant Pathol. 2009 Nov;10(6):837-42. doi: 10.1111/j.1364-3703.2009.00577.x. Mol Plant Pathol. 2009. PMID: 19849789 Free PMC article.
-
Closing the circle on the discovery of genes encoding Hrp regulon members and type III secretion system effectors in the genomes of three model Pseudomonas syringae strains.Mol Plant Microbe Interact. 2006 Nov;19(11):1151-8. doi: 10.1094/MPMI-19-1151. Mol Plant Microbe Interact. 2006. PMID: 17073298 Review.
-
Regulation of the type III secretion system in phytopathogenic bacteria.Mol Plant Microbe Interact. 2006 Nov;19(11):1159-66. doi: 10.1094/MPMI-19-1159. Mol Plant Microbe Interact. 2006. PMID: 17073299 Review.
Cited by
-
Noncanonical calcium binding motif controls folding of HopQ1, a Pseudomonas syringae type III secretion effector, in a pH-dependent manner.Sci Rep. 2024 Dec 30;14(1):31796. doi: 10.1038/s41598-024-82848-z. Sci Rep. 2024. PMID: 39738557 Free PMC article.
-
Formation of HopQ1:14-3-3 complex in the host cytoplasm modulates nuclear import rate of Pseudomonas syringae effector in Nicotiana benthamiana cells.Front Plant Sci. 2024 Mar 4;15:1335830. doi: 10.3389/fpls.2024.1335830. eCollection 2024. Front Plant Sci. 2024. PMID: 38501137 Free PMC article.
-
A Bacterial Effector Mimics a Host HSP90 Client to Undermine Immunity.Cell. 2019 Sep 19;179(1):205-218.e21. doi: 10.1016/j.cell.2019.08.020. Epub 2019 Sep 12. Cell. 2019. PMID: 31522888 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous