

Invariant Natural Killer T Cell Subsets-More Than Just Developmental Intermediates
- PMID: 29973936
- PMCID: PMC6019445
- DOI: 10.3389/fimmu.2018.01393
Invariant Natural Killer T Cell Subsets-More Than Just Developmental Intermediates
Abstract
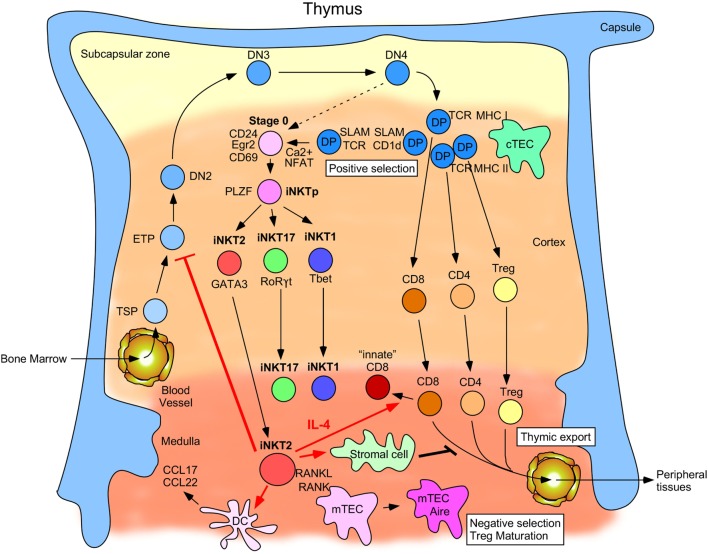
Invariant natural killer T (iNKT) cells are a CD1d-restricted T cell population that can respond to lipid antigenic stimulation within minutes by secreting a wide variety of cytokines. This broad functional scope has placed iNKT cells at the frontlines of many kinds of immune responses. Although the diverse functional capacities of iNKT cells have long been acknowledged, only recently have distinct iNKT cell subsets, each with a marked functional predisposition, been appreciated. Furthermore, the subsets can frequently occupy distinct niches in different tissues and sometimes establish long-term tissue residency where they can impact homeostasis and respond quickly when they sense perturbations. In this review, we discuss the developmental origins of the iNKT cell subsets, their localization patterns, and detail what is known about how different subsets specifically influence their surroundings in conditions of steady and diseased states.
Keywords: cytokine secretion; development; homeostasis; invariant natural killer T cells; subsets.
Figures
Similar articles
-
Serpinb1a Is Dispensable for the Development and Cytokine Response of Invariant Natural Killer T Cell Subsets.Front Immunol. 2020 Nov 11;11:562587. doi: 10.3389/fimmu.2020.562587. eCollection 2020. Front Immunol. 2020. PMID: 33262755 Free PMC article.
-
How Lipid-Specific T Cells Become Effectors: The Differentiation of iNKT Subsets.Front Immunol. 2018 Jun 26;9:1450. doi: 10.3389/fimmu.2018.01450. eCollection 2018. Front Immunol. 2018. PMID: 29997620 Free PMC article. Review.
-
Distinct and overlapping effector functions of expanded human CD4+, CD8α+ and CD4-CD8α- invariant natural killer T cells.PLoS One. 2011;6(12):e28648. doi: 10.1371/journal.pone.0028648. Epub 2011 Dec 12. PLoS One. 2011. PMID: 22174854 Free PMC article.
-
Functions of CD1d-Restricted Invariant Natural Killer T Cells in Antimicrobial Immunity and Potential Applications for Infection Control.Front Immunol. 2018 Jun 6;9:1266. doi: 10.3389/fimmu.2018.01266. eCollection 2018. Front Immunol. 2018. PMID: 29928278 Free PMC article. Review.
-
Strategy of lipid recognition by invariant natural killer T cells: 'one for all and all for one'.Immunology. 2012 Jul;136(3):273-82. doi: 10.1111/j.1365-2567.2012.03580.x. Immunology. 2012. PMID: 22671023 Free PMC article. Review.
Cited by
-
Natural Killer T Cell Diversity and Immunotherapy.Cancers (Basel). 2023 Dec 7;15(24):5737. doi: 10.3390/cancers15245737. Cancers (Basel). 2023. PMID: 38136283 Free PMC article. Review.
-
Does exercise attenuate age- and disease-associated dysfunction in unconventional T cells? Shining a light on overlooked cells in exercise immunology.Eur J Appl Physiol. 2021 Jul;121(7):1815-1834. doi: 10.1007/s00421-021-04679-4. Epub 2021 Apr 6. Eur J Appl Physiol. 2021. PMID: 33822261 Review.
-
Current insights in mouse iNKT and MAIT cell development using single cell transcriptomics data.Semin Immunol. 2022 Mar;60:101658. doi: 10.1016/j.smim.2022.101658. Epub 2022 Sep 28. Semin Immunol. 2022. PMID: 36182863 Free PMC article. Review.
-
Type 2 hypersensitivity disorders, including systemic lupus erythematosus, Sjögren's syndrome, Graves' disease, myasthenia gravis, immune thrombocytopenia, autoimmune hemolytic anemia, dermatomyositis, and graft-versus-host disease, are THαβ-dominant autoimmune diseases.Virulence. 2024 Dec;15(1):2404225. doi: 10.1080/21505594.2024.2404225. Epub 2024 Sep 16. Virulence. 2024. PMID: 39267271 Free PMC article. Review.
-
Ezh2 Shapes T Cell Plasticity to Drive Atherosclerosis.Circulation. 2025 May 13;151(19):1391-1408. doi: 10.1161/CIRCULATIONAHA.124.072384. Epub 2025 Feb 7. Circulation. 2025. PMID: 39917842 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources