

Genetic basis and evolution of rapid cycling in railway populations of tetraploid Arabidopsis arenosa
- PMID: 29975688
- PMCID: PMC6049958
- DOI: 10.1371/journal.pgen.1007510
Genetic basis and evolution of rapid cycling in railway populations of tetraploid Arabidopsis arenosa
Abstract
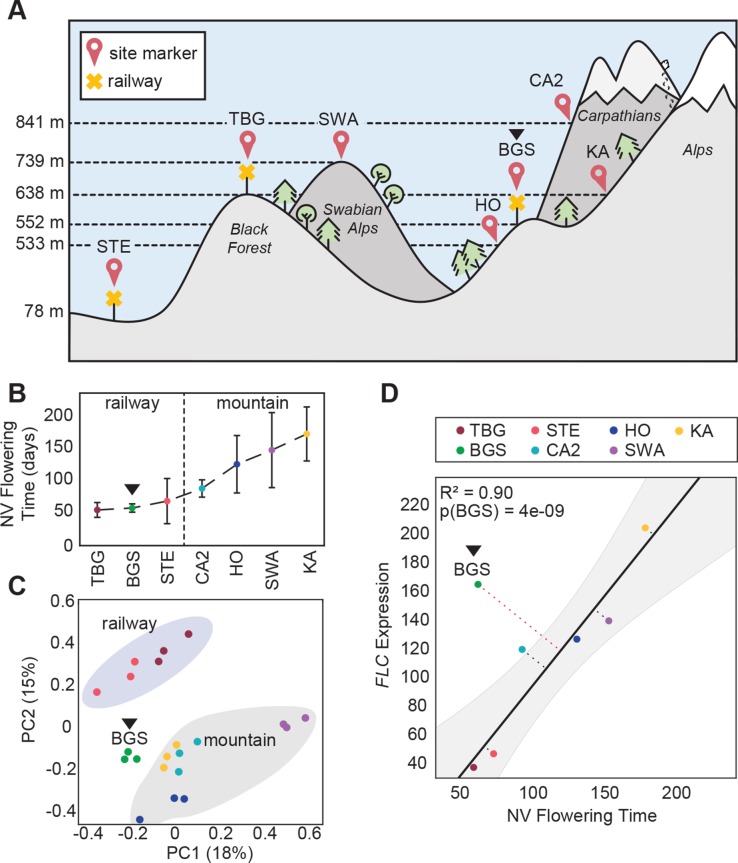
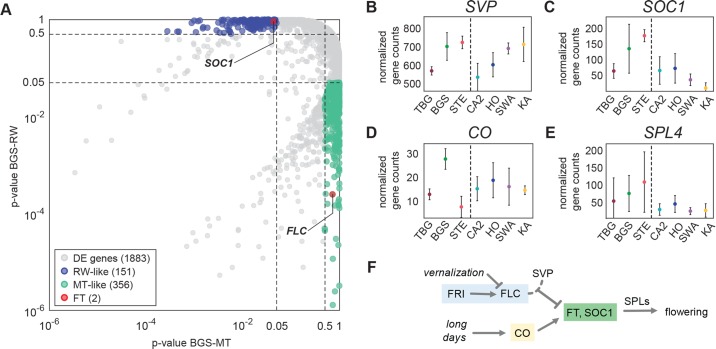
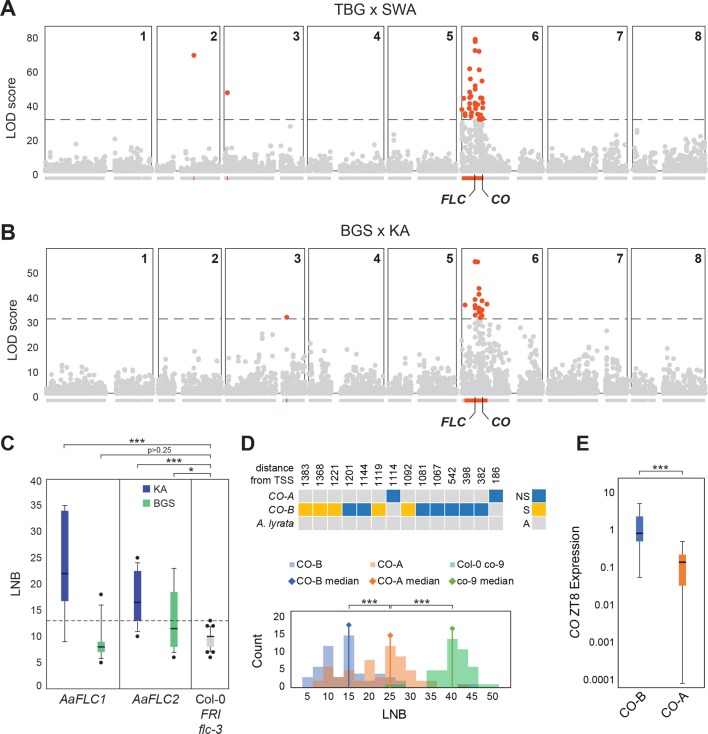
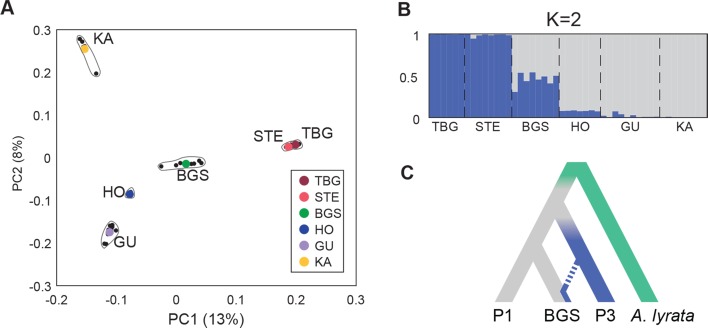
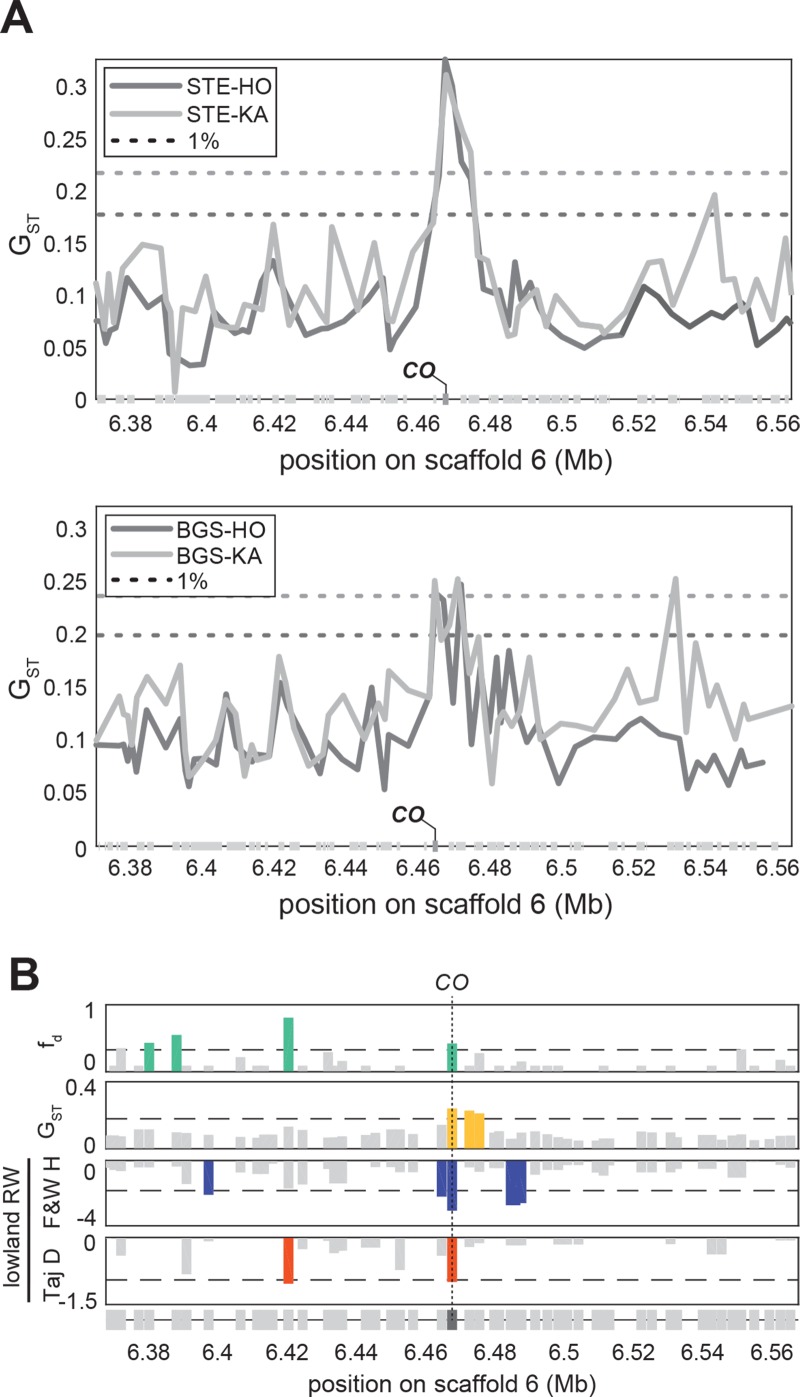
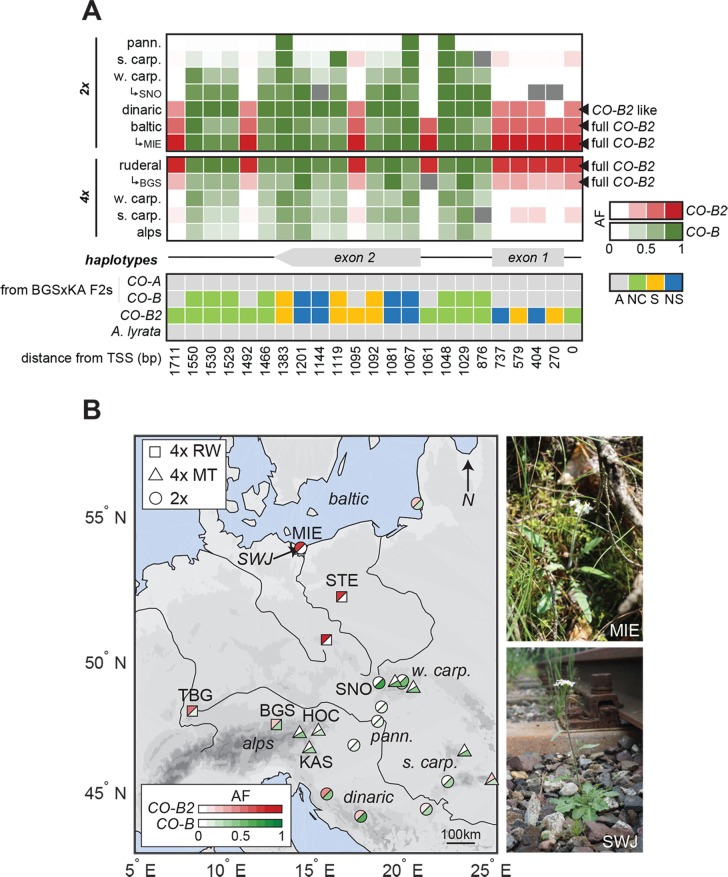
Spatially structured plant populations with diverse adaptations provide powerful models to investigate evolution. Human-generated ruderal habitats are abundant and low-competition, but are challenging for plants not adapted to them. Ruderal habitats also sometimes form networked corridors (e.g. roadsides and railways) that allow rapid long-distance spread of successfully adapted variants. Here we use transcriptomic and genomic analyses, coupled with genetic mapping and transgenic follow-up, to understand the evolution of rapid cycling during adaptation to railway sites in autotetraploid Arabidopsis arenosa. We focus mostly on a hybrid population that is likely a secondary colonist of a railway site. These mountain railway plants are phenotypically similar to their cosmopolitan cousins. We thus hypothesized that colonization primarily involved the flow of adaptive alleles from the cosmopolitan railway variant. But our data shows that it is not that simple: while there is evidence of selection having acted on introgressed alleles, selection also acted on rare standing variation, and new mutations may also contribute. Among the genes we show have allelic divergence with functional relevance to flowering time are known regulators of flowering, including FLC and CONSTANS. Prior implications of these genes in weediness and rapid cycling supports the idea that these are "evolutionary hotspots" for these traits. We also find that one of two alleles of CONSTANS under selection in the secondary colonist was selected from rare standing variation in mountain populations, while the other was introgressed from the cosmopolitan railway populations. The latter allele likely arose in diploid populations over 700km away, highlighting how ruderal populations could act as allele conduits and thus influence local adaptation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Hall MC, Willis JH. Divergent selection on flowering time contributes to local adaptation in Mimulus guttatus populations. Evolution. 2006. December;60(12):2466–77. - PubMed
-
- Fox GA. Drought and the evolution of flowering time in desert annuals. American Journal of Botany. 1990. 77: 1508–18.
-
- Sherrard ME, Maherali H. The adaptive significance of drought escape in Avena barbata, an annual grass. Evolution (N Y). 2006. December;60(12):2478–89. - PubMed
-
- Mckay JK, Richards JH, Mitchell-Olds T. Genetics of drought adaptation in Arabidopsis thaliana: I. Pleiotropy contributes to genetic correlations among ecological traits. Mol Ecol. 2003. May;12(5):1137–51. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
