

Sad1 Spatiotemporally Regulates Kinetochore Clustering To Ensure High-Fidelity Chromosome Segregation in the Human Fungal Pathogen Cryptococcus neoformans
- PMID: 29976642
- PMCID: PMC6034078
- DOI: 10.1128/mSphere.00190-18
Sad1 Spatiotemporally Regulates Kinetochore Clustering To Ensure High-Fidelity Chromosome Segregation in the Human Fungal Pathogen Cryptococcus neoformans
Abstract
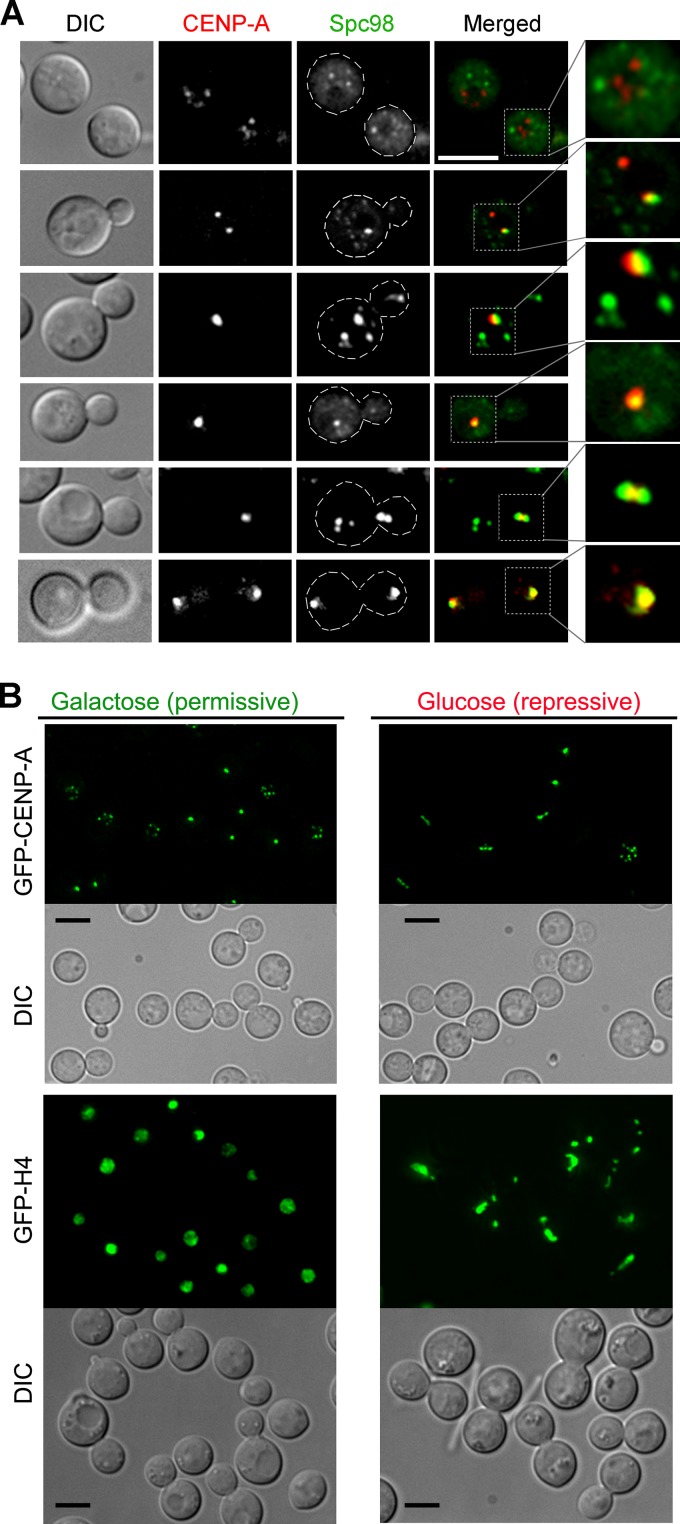
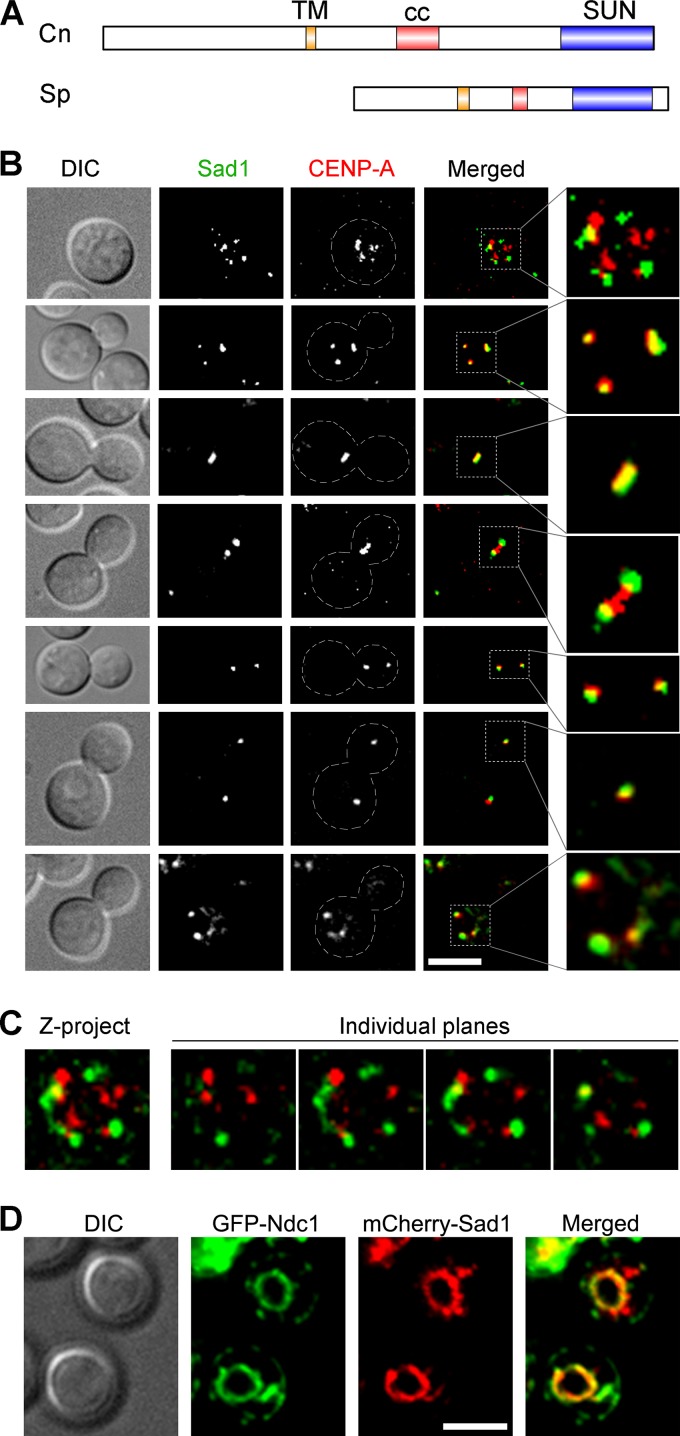
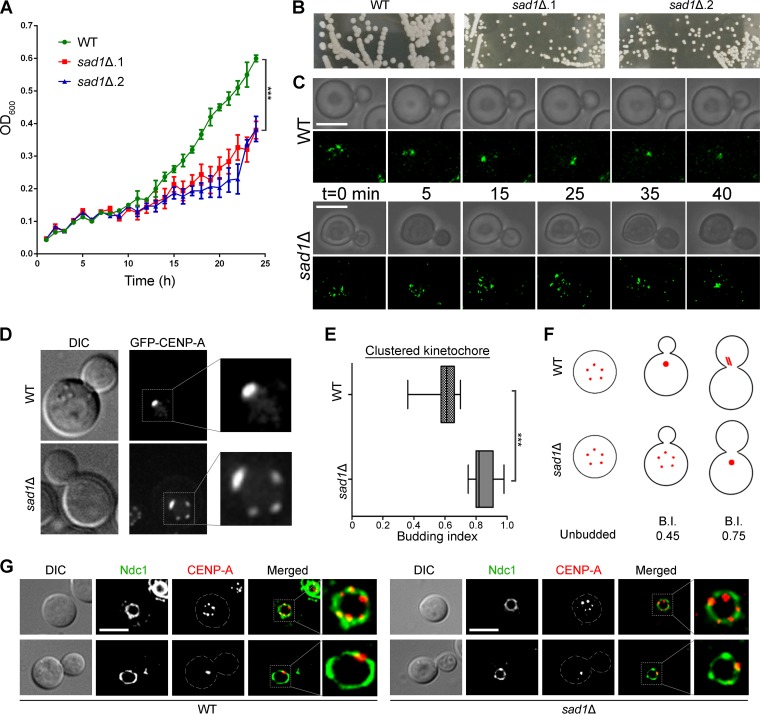
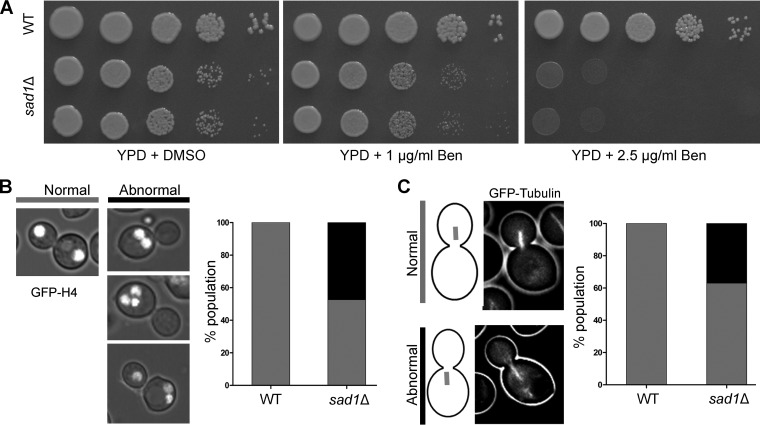
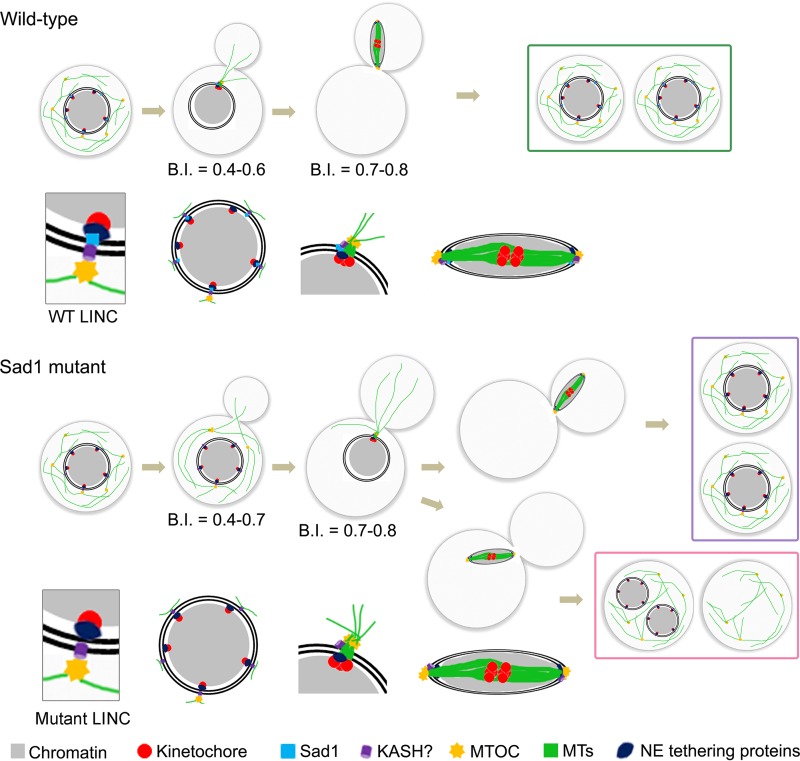
Kinetochore clustering, frequently observed in yeasts, plays a key role in genome organization and chromosome segregation. In the absence of the metaphase plate arrangement, kinetochore clustering in yeast species is believed to facilitate timely kinetochore-microtubule interactions to achieve bivalent attachments of chromosomes during metaphase. The factors determining the dynamics of kinetochore clustering remain largely unknown. We previously reported that kinetochores oscillate between an unclustered and a clustered state during the mitotic cell cycle in the basidiomycetous yeast Cryptococcus neoformans Based on tubulin localization patterns, while kinetochore clustering appears to be microtubule dependent, an indirect interaction of microtubules with kinetochores is expected in C. neoformans In this study, we sought to examine possible roles of the SUN-KASH protein complex, known to form a bridge across the nuclear envelope, in regulating kinetochore clustering in C. neoformans We show that the SUN domain protein Sad1 localizes close to kinetochores in interphase as well as in mitotic cells. Sad1 is nonessential for viability in C. neoformans but is required for proper growth and high-fidelity chromosome segregation. Further, we demonstrate that the onset of kinetochore clustering is significantly delayed in cells lacking Sad1 compared to wild-type cells. Taken together, this study identifies a novel role of the SUN domain protein Sad1 in spatiotemporal regulation of kinetochore clustering during the mitotic cell cycle in C. neoformansIMPORTANCE The linker of nucleoskeleton and cytoskeleton (LINC) complex is present in fungi, animals, and plants. It performs diverse functions in animals, and its role(s) have recently been explored in plants. In ascomycetous yeast species, the role of the LINC complex in spindle pole body function and telomere clustering during meiosis has been determined. However, nothing is known about the LINC complex in the fungal phylum of Basidiomycota. In this study, we identified the role of the LINC complex in kinetochore dynamics as well as in nuclear migration in a basidiomycetous yeast, Cryptococcus neoformans, a human pathogen. Unlike most other yeast species, kinetochores remain unclustered during interphase but gradually cluster during mitosis in C. neoformans We report that the LINC complex is required for timely onset of kinetochore clustering and high-fidelity chromosome segregation in C. neoformans Thus, our study identifies a novel factor required for kinetochore clustering during mitosis in yeast species.
Keywords: CENP-A; LINC complex; microtubule organizing center; mitotic spindle.
Copyright © 2018 Yadav and Sanyal.
Figures
References
-
- Jin QW, Fuchs J, Loidl J. 2000. Centromere clustering is a major determinant of yeast interphase nuclear organization. J Cell Sci 113:1903–1912. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
