

Reprogramming of basic metabolic pathways in microbial sepsis: therapeutic targets at last?
- PMID: 29976786
- PMCID: PMC6079534
- DOI: 10.15252/emmm.201708712
Reprogramming of basic metabolic pathways in microbial sepsis: therapeutic targets at last?
Abstract
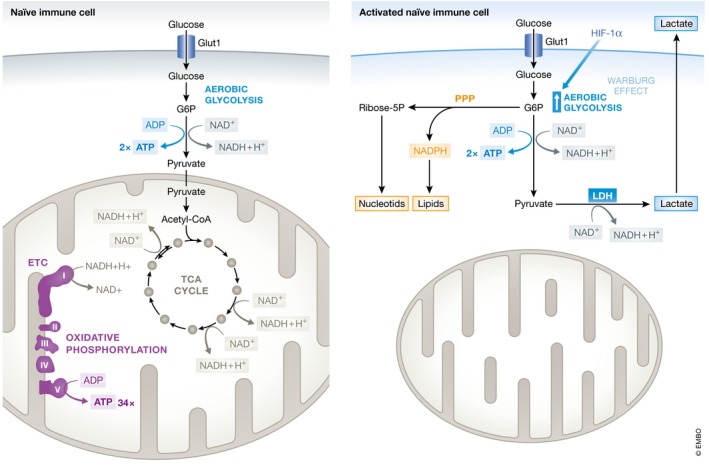
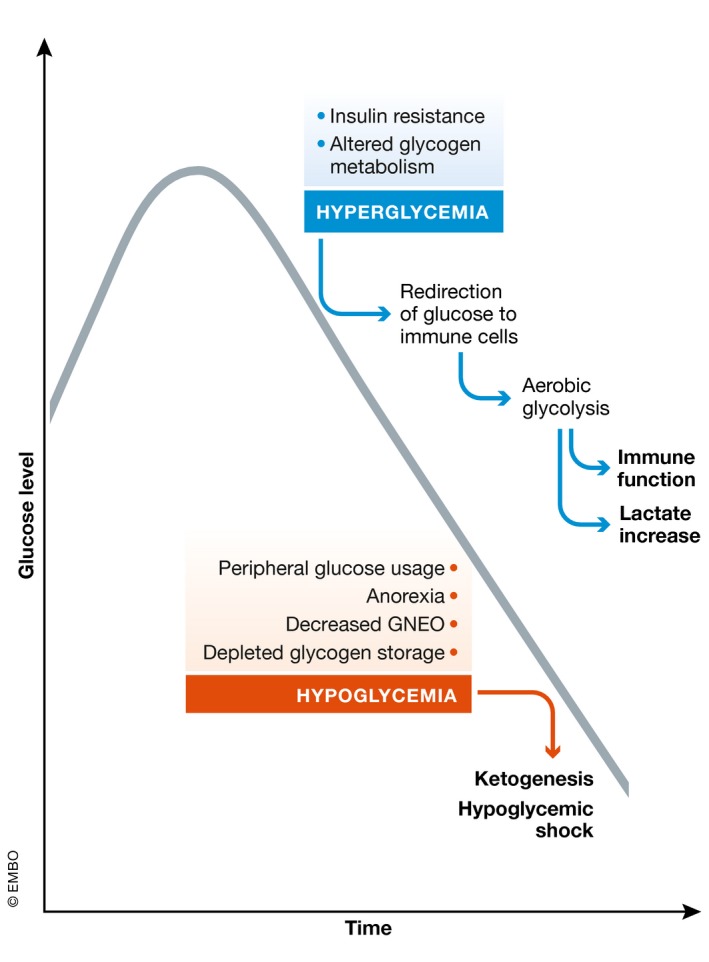
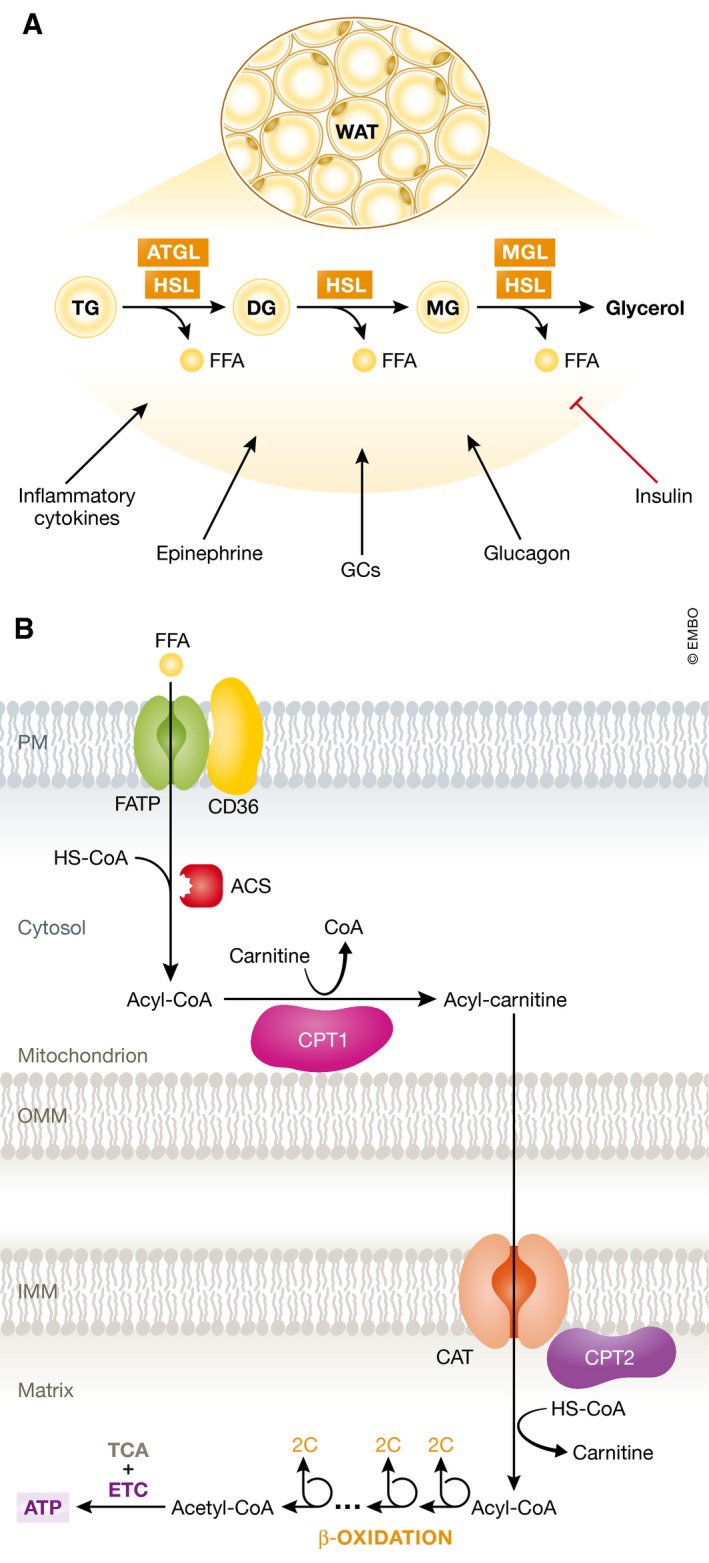
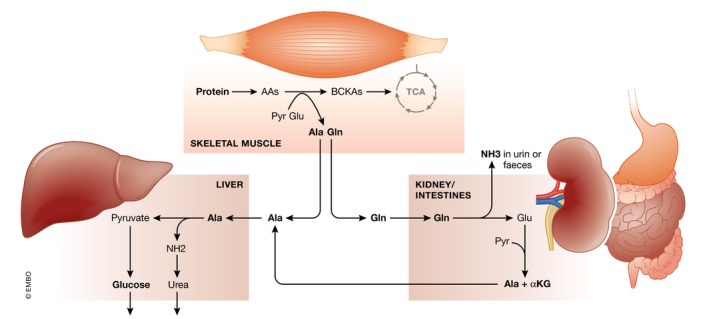
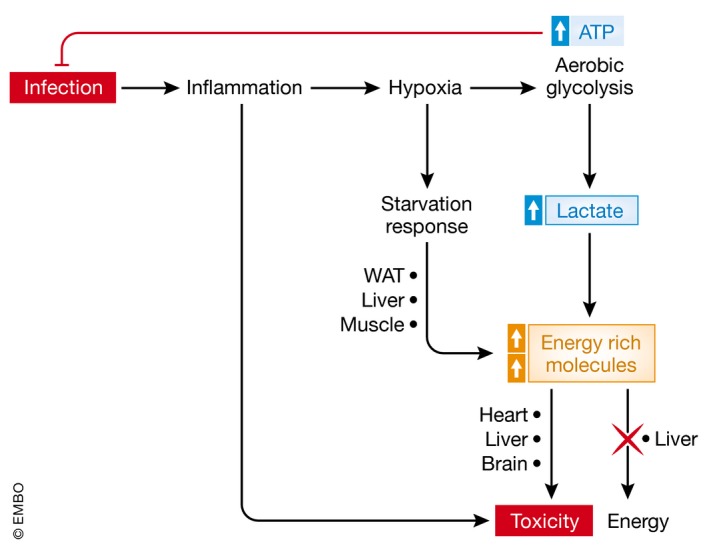
Sepsis is a highly lethal and urgent unmet medical need. It is the result of a complex interplay of several pathways, including inflammation, immune activation, hypoxia, and metabolic reprogramming. Specifically, the regulation and the impact of the latter have become better understood in which the highly catabolic status during sepsis and its similarity with starvation responses appear to be essential in the poor prognosis in sepsis. It seems logical that new interventions based on the recognition of new therapeutic targets in the key metabolic pathways should be developed and may have a good chance to penetrate to the bedside. In this review, we concentrate on the pathological changes in metabolism, observed during sepsis, and the presumed underlying mechanisms, with a focus on the level of the organism and the interplay between different organ systems.
Keywords: hypoxia; inflammation; interventions; metabolic reprogramming; sepsis.
© 2018 The Authors. Published under the terms of the CC BY 4.0 license.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
