

Zebrafish mutants and TEAD reporters reveal essential functions for Yap and Taz in posterior cardinal vein development
- PMID: 29976931
- PMCID: PMC6033906
- DOI: 10.1038/s41598-018-27657-x
Zebrafish mutants and TEAD reporters reveal essential functions for Yap and Taz in posterior cardinal vein development
Abstract
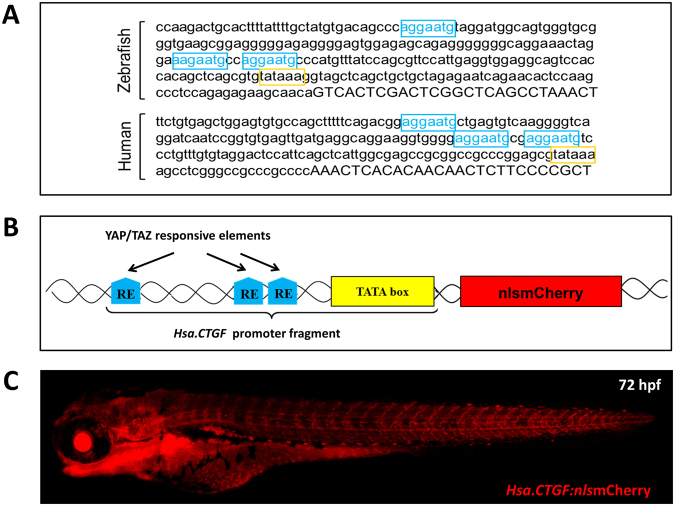
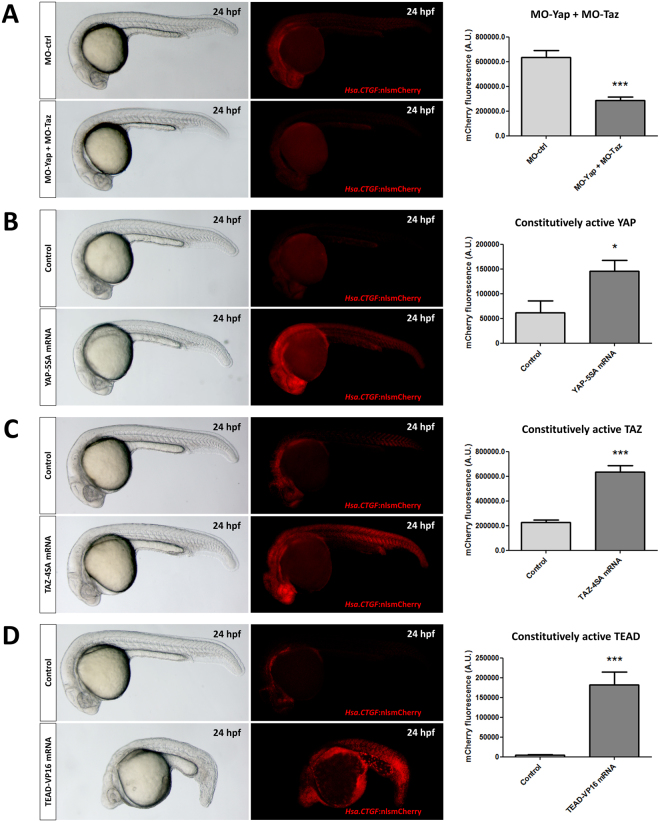
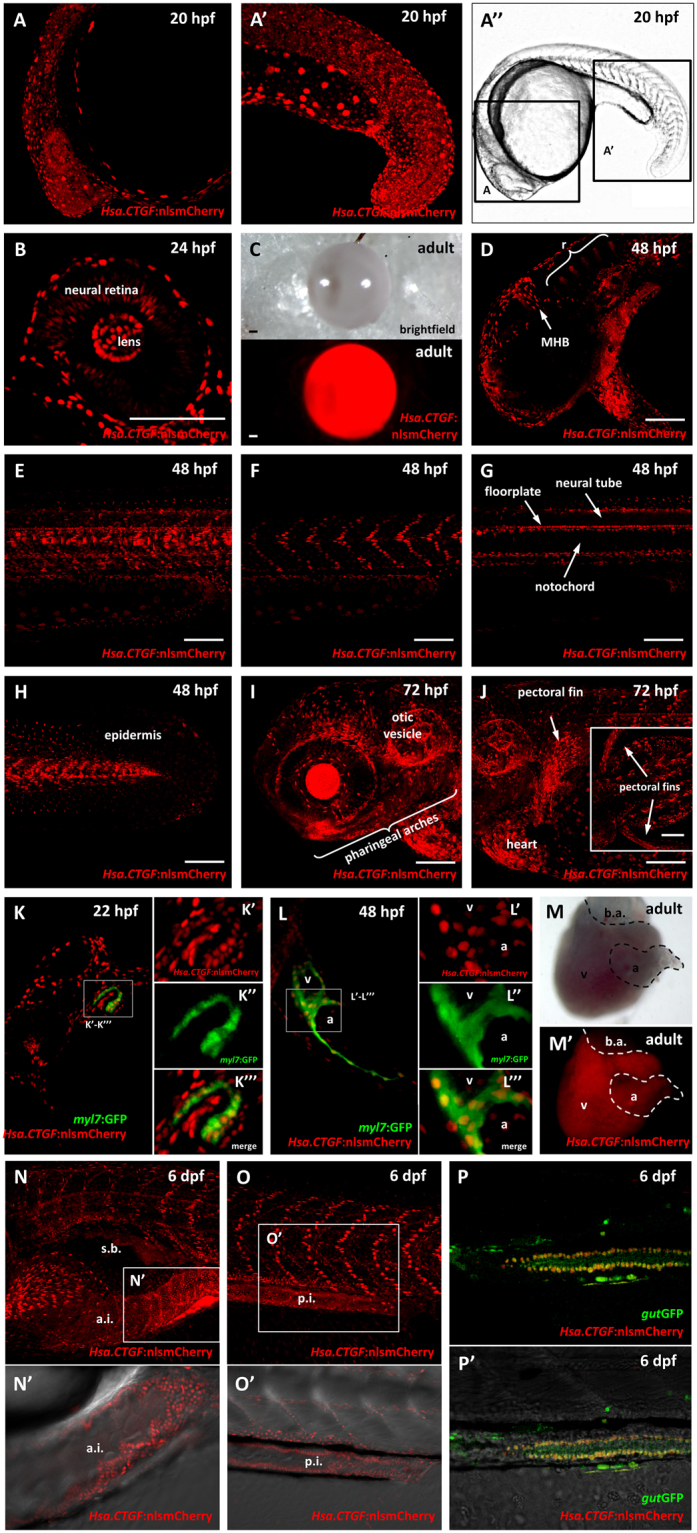
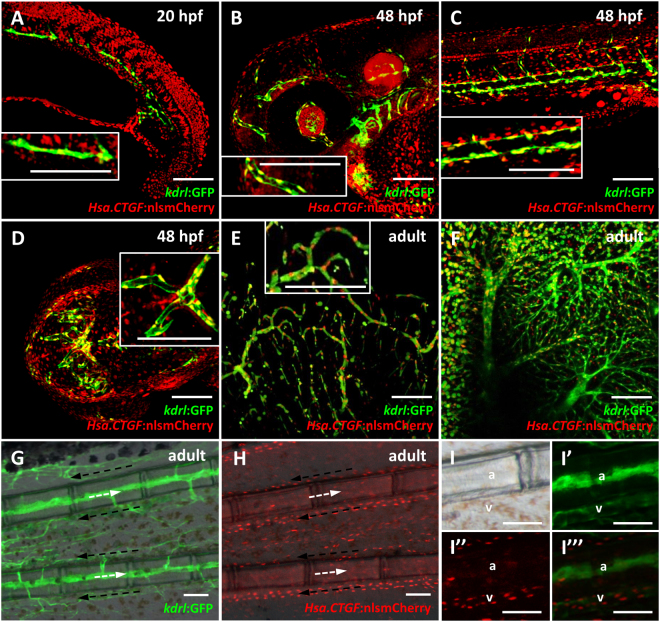
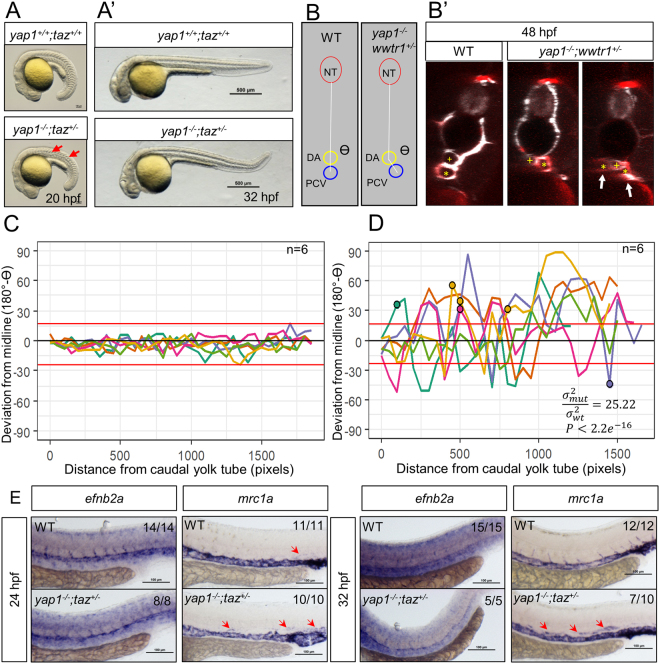
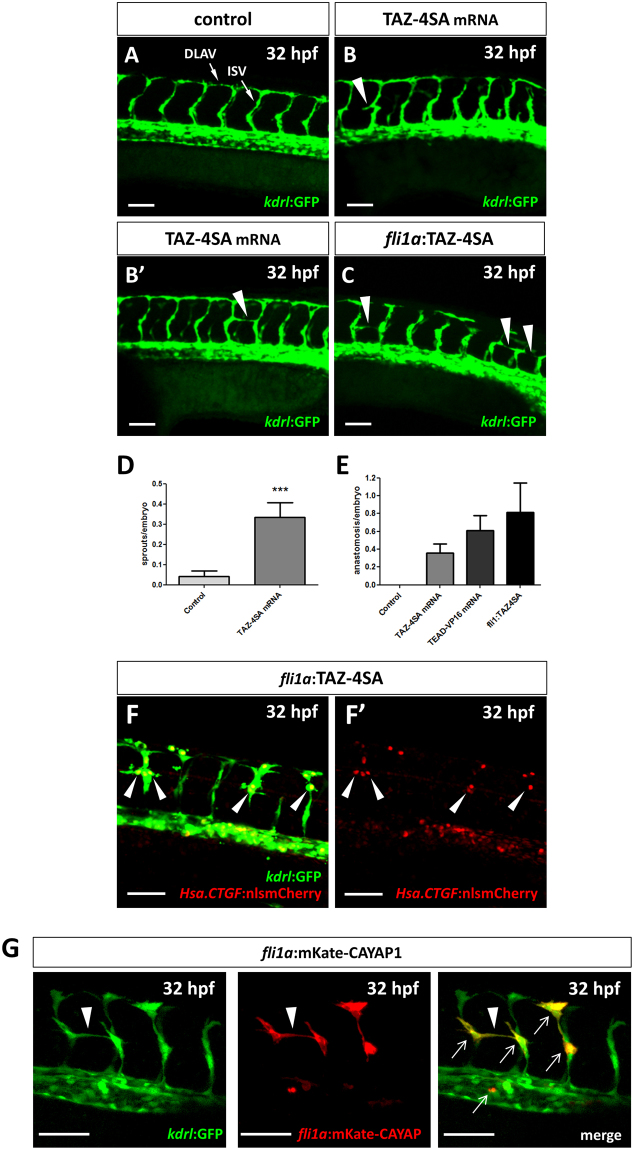
As effectors of the Hippo signaling cascade, YAP1 and TAZ are transcriptional regulators playing important roles in development, tissue homeostasis and cancer. A number of different cues, including mechanotransduction of extracellular stimuli, adhesion molecules, oncogenic signaling and metabolism modulate YAP1/TAZ nucleo-cytoplasmic shuttling. In the nucleus, YAP1/TAZ tether with the DNA binding proteins TEADs, to activate the expression of target genes that regulate proliferation, migration, cell plasticity, and cell fate. Based on responsive elements present in the human and zebrafish promoters of the YAP1/TAZ target gene CTGF, we established zebrafish fluorescent transgenic reporter lines of Yap1/Taz activity. These reporter lines provide an in vivo view of Yap1/Taz activity during development and adulthood at the whole organism level. Transgene expression was detected in many larval tissues including the otic vesicles, heart, pharyngeal arches, muscles and brain and is prominent in endothelial cells. Analysis of vascular development in yap1/taz zebrafish mutants revealed specific defects in posterior cardinal vein (PCV) formation, with altered expression of arterial/venous markers. The overactivation of Yap1/Taz in endothelial cells was sufficient to promote an aberrant vessel sprouting phenotype. Our findings confirm and extend the emerging role of Yap1/Taz in vascular development including angiogenesis.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
