

Spontaneous reactivation of latent HIV-1 promoters is linked to the cell cycle as revealed by a genetic-insulators-containing dual-fluorescence HIV-1-based vector
- PMID: 29977044
- PMCID: PMC6033903
- DOI: 10.1038/s41598-018-28161-y
Spontaneous reactivation of latent HIV-1 promoters is linked to the cell cycle as revealed by a genetic-insulators-containing dual-fluorescence HIV-1-based vector
Abstract
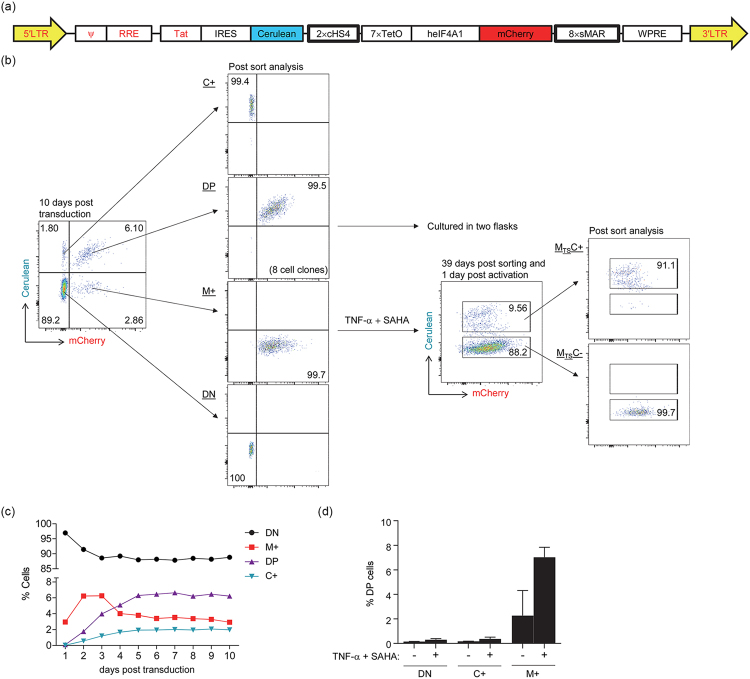
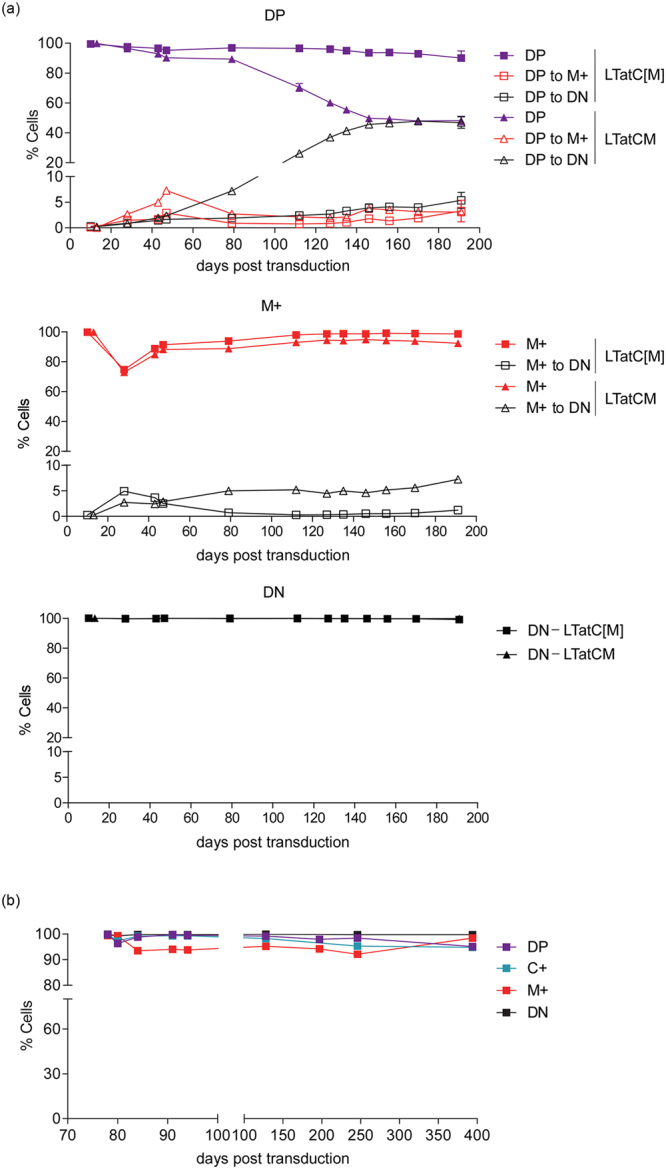
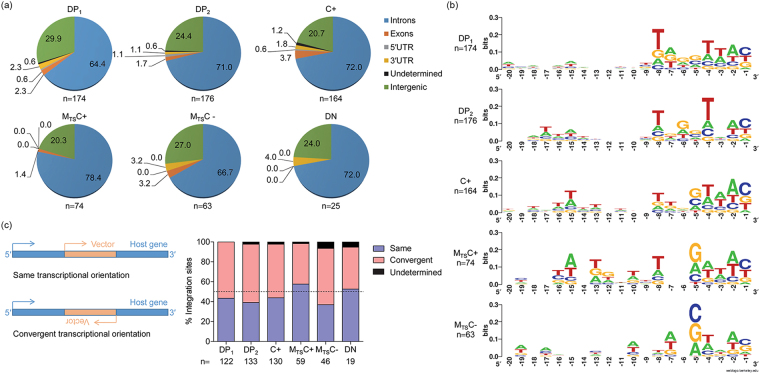
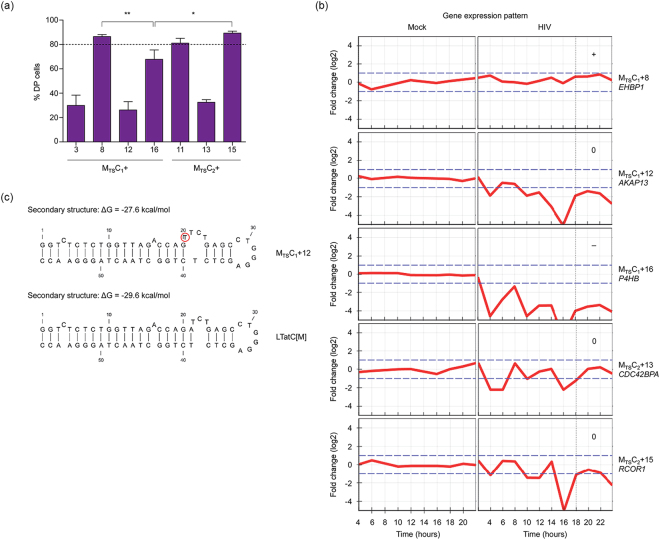
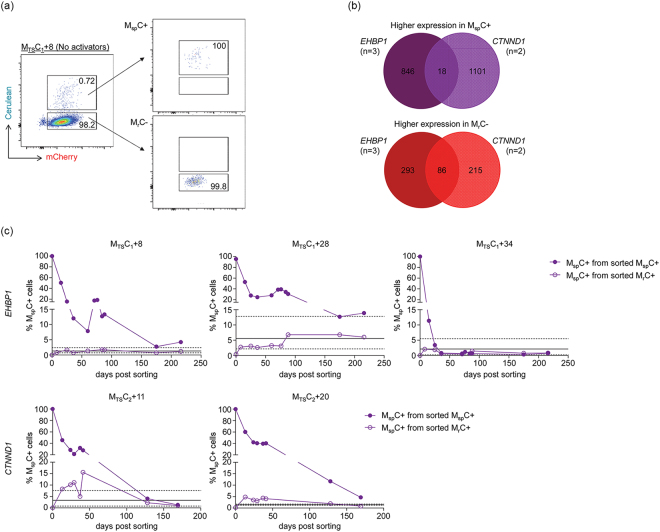
Long-lived latently HIV-1-infected cells represent a barrier to cure. We developed a dual-fluorescence HIV-1-based vector containing a pair of genetic insulators flanking a constitutive fluorescent reporter gene to study HIV-1 latency. The protective effects of these genetic insulators are demonstrated through long-term (up to 394 days) stable fluorescence profiles in transduced SUP-T1 cells. Analysis of 1,941 vector integration sites confirmed reproduction of HIV-1 integration patterns. We sorted monoclonal cells representing latent HIV-1 infections and found that both vector integration sites and integrity of the vector genomes influence the reactivation potentials of latent HIV-1 promoters. Interestingly, some latent monoclonal cells exhibited a small cell subpopulation with a spontaneously reactivated HIV-1 promoter. Higher expression levels of genes involved in cell cycle progression are observed in these cell subpopulations compared to their counterparts with HIV-1 promoters that remained latent. Consistently, larger fractions of spontaneously reactivated cells are in the S and G2 phases of the cell cycle. Furthermore, genistein and nocodazole treatments of these cell clones, which halted cells in the G2 phase, resulted in a 1.4-2.9-fold increase in spontaneous reactivation. Taken together, our HIV-1 latency model reveals that the spontaneous reactivation of latent HIV-1 promoters is linked to the cell cycle.
Conflict of interest statement
K.J.M. has received travel grants and honoraria from Gilead Sciences, Roche Diagnostics, Tibotec, Bristol-Myers Squibb, and Abbott; the University of Zurich has received research grants from Gilead, Roche, and Merck Sharp & Dohme for studies that K.J.M. serves as principal investigator, and advisory board honoraria from Gilead Sciences. All other authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
