

Evolutionary trade-offs associated with loss of PmrB function in host-adapted Pseudomonas aeruginosa
- PMID: 29980663
- PMCID: PMC6035264
- DOI: 10.1038/s41467-018-04996-x
Evolutionary trade-offs associated with loss of PmrB function in host-adapted Pseudomonas aeruginosa
Abstract
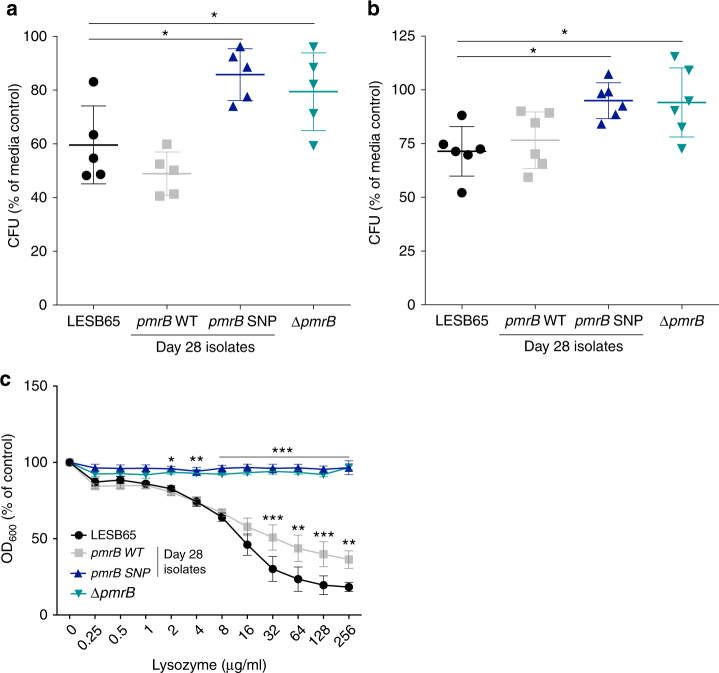
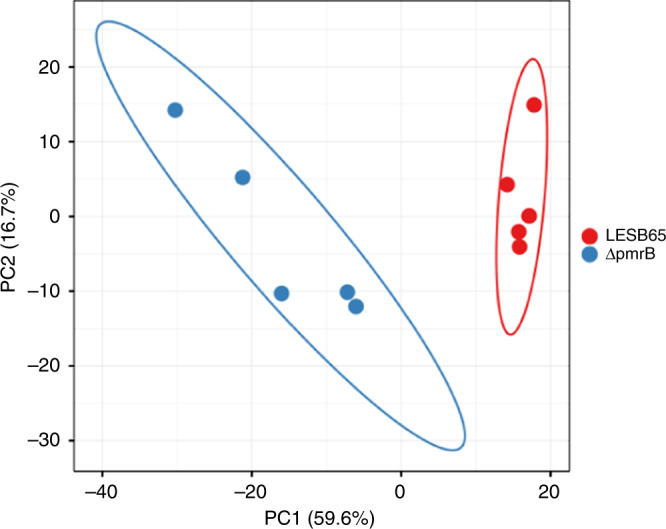
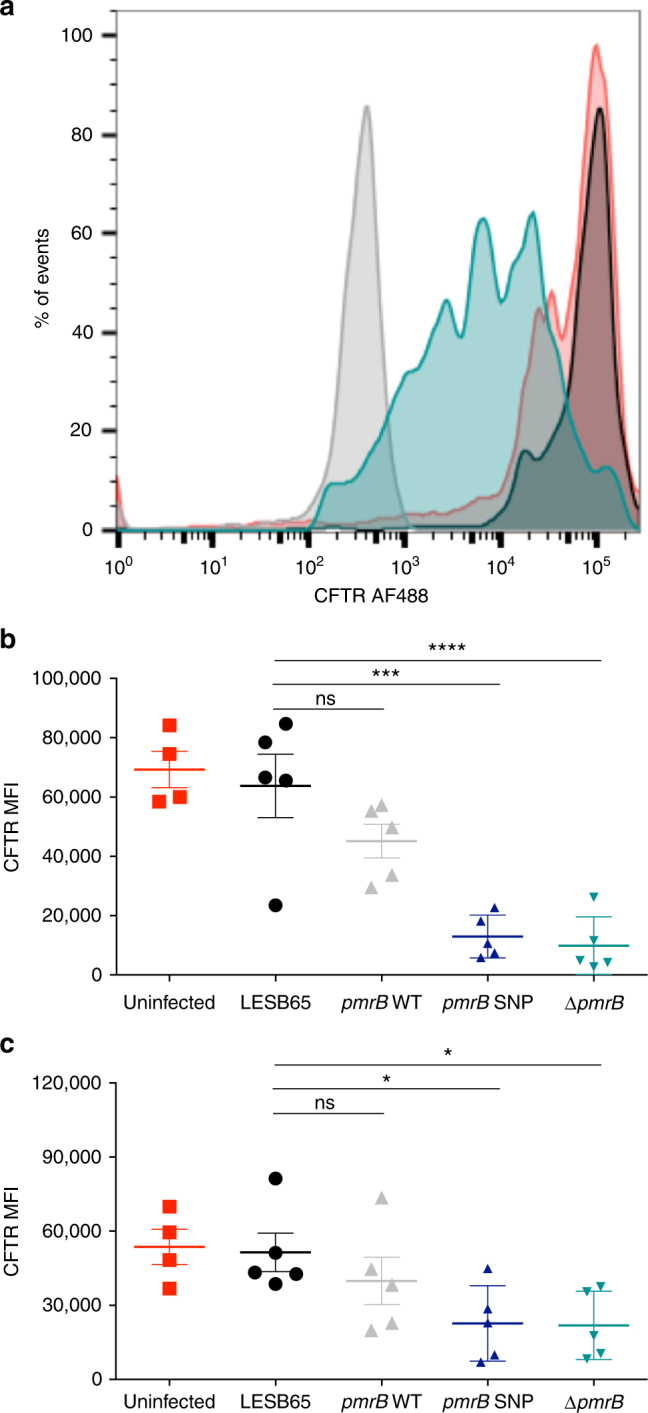
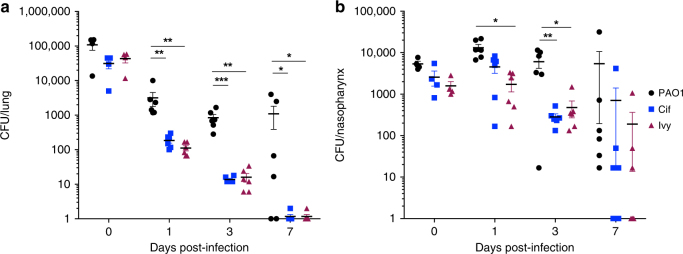
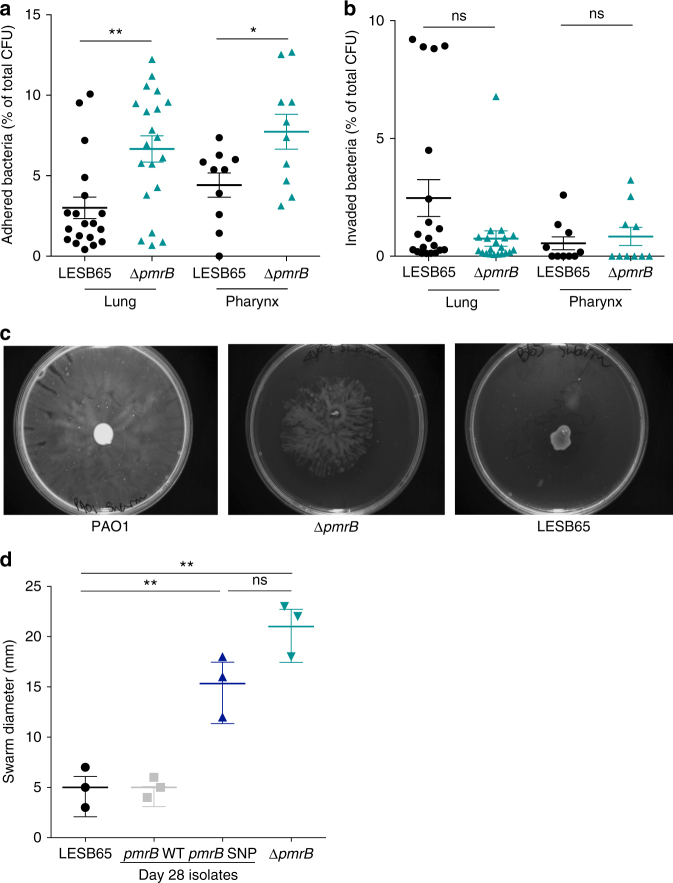
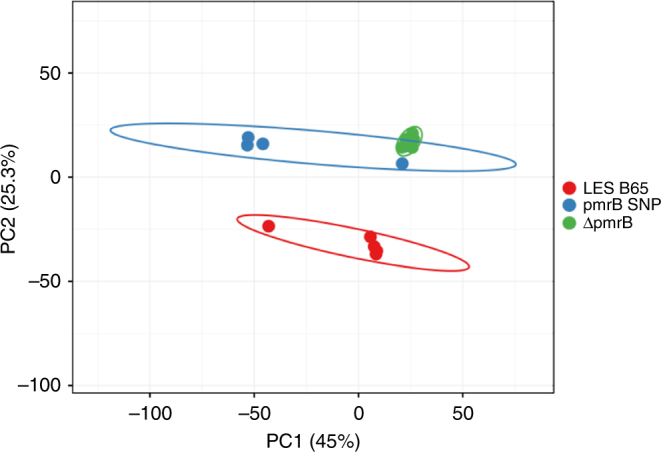
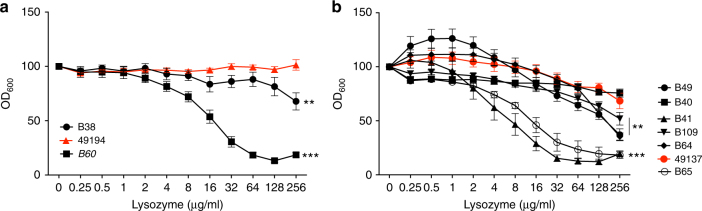
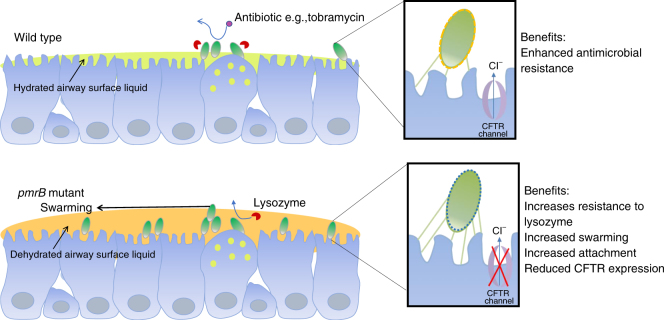
Pseudomonas aeruginosa colonises the upper airway of cystic fibrosis (CF) patients, providing a reservoir of host-adapted genotypes that subsequently establish chronic lung infection. We previously experimentally-evolved P. aeruginosa in a murine model of respiratory tract infection and observed early-acquired mutations in pmrB, encoding the sensor kinase of a two-component system that promoted establishment and persistence of infection. Here, using proteomics, we show downregulation of proteins involved in LPS biosynthesis, antimicrobial resistance and phenazine production in pmrB mutants, and upregulation of proteins involved in adherence, lysozyme resistance and inhibition of the chloride ion channel CFTR, relative to wild-type strain LESB65. Accordingly, pmrB mutants are susceptible to antibiotic treatment but show enhanced adherence to airway epithelial cells, resistance to lysozyme treatment, and downregulate host CFTR expression. We propose that P. aeruginosa pmrB mutations in CF patients are subject to an evolutionary trade-off, leading to enhanced colonisation potential, CFTR inhibition, and resistance to host defences, but also to increased susceptibility to antibiotics.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
