

Histone H1 acetylation at lysine 85 regulates chromatin condensation and genome stability upon DNA damage
- PMID: 29982688
- PMCID: PMC6125638
- DOI: 10.1093/nar/gky568
Histone H1 acetylation at lysine 85 regulates chromatin condensation and genome stability upon DNA damage
Abstract
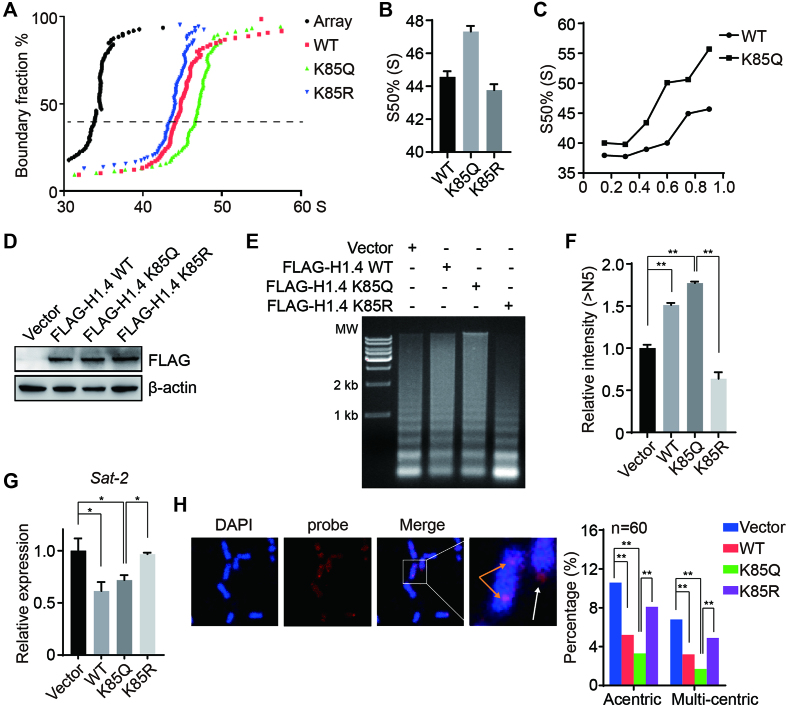
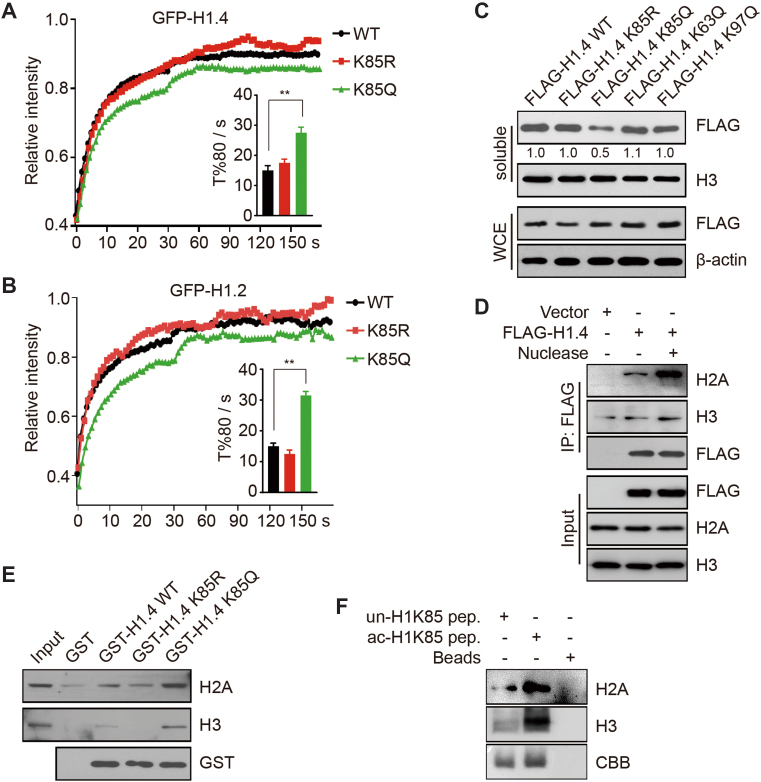
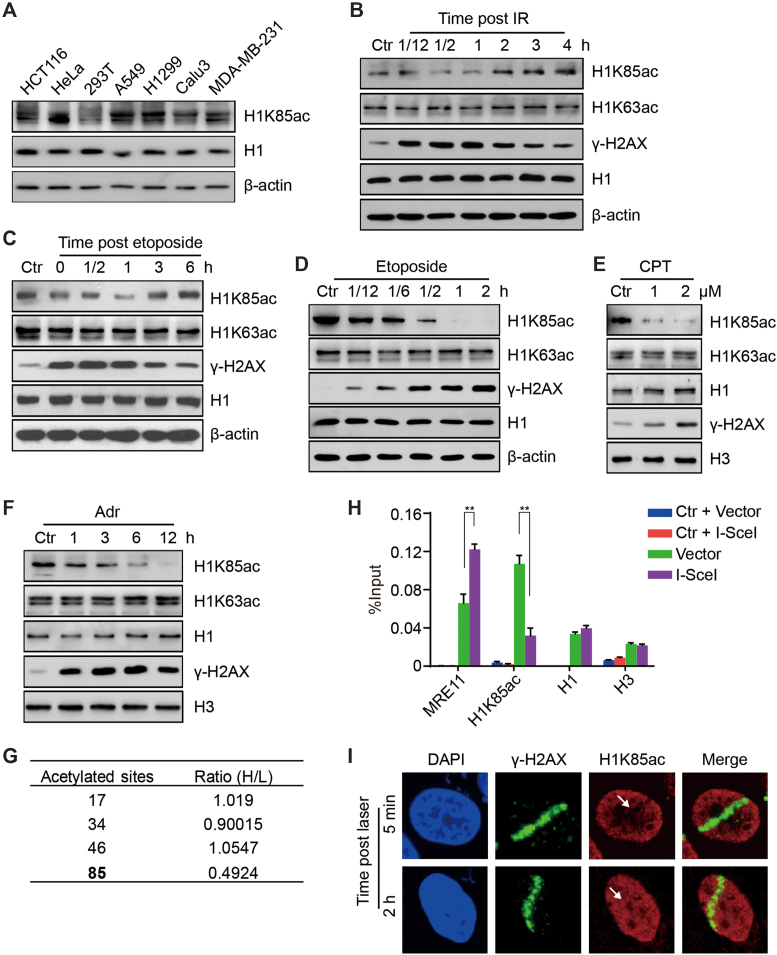
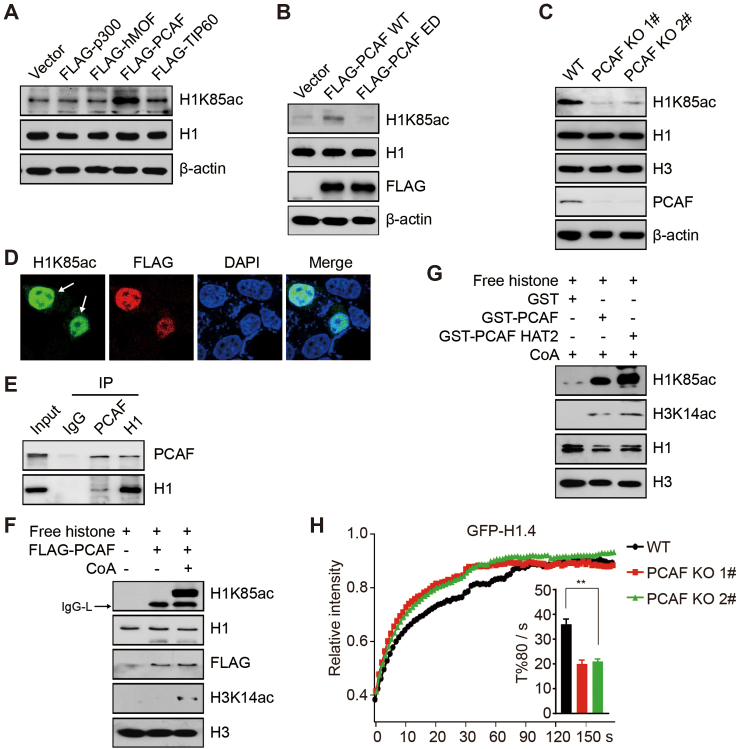
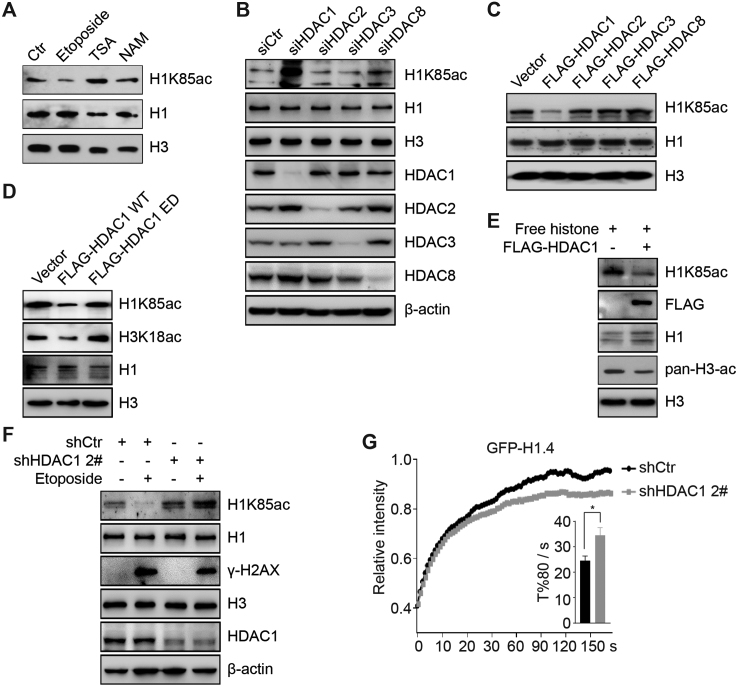
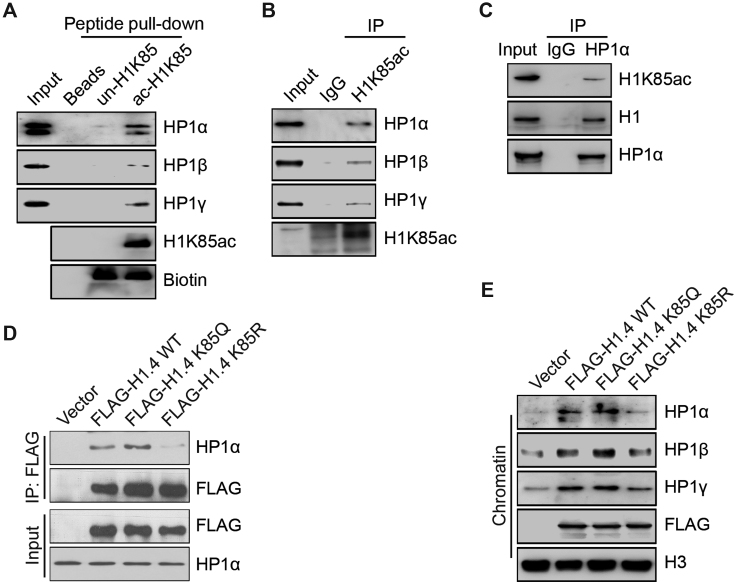
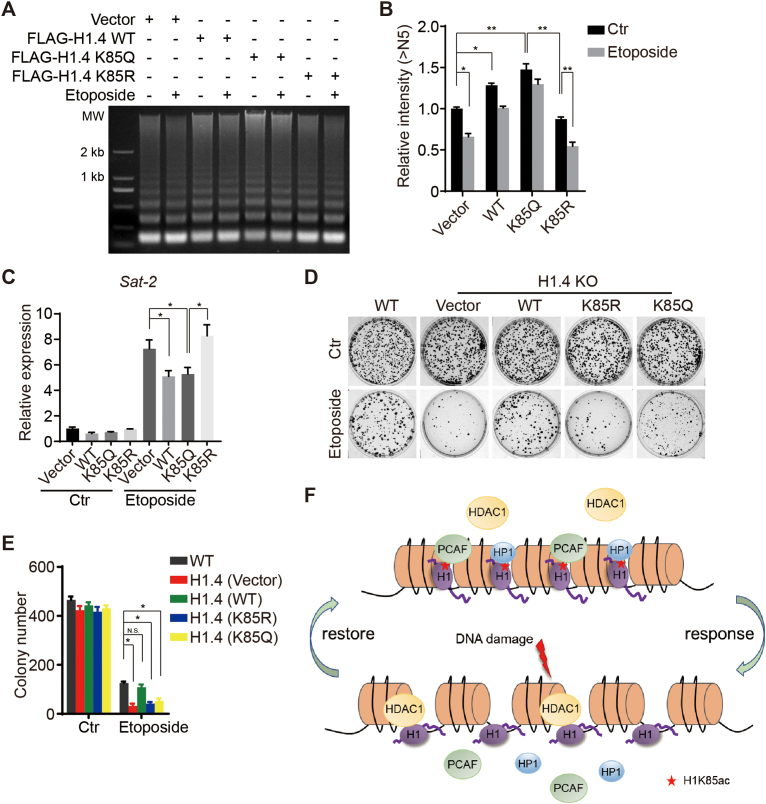
Linker histone H1 has a key role in maintaining higher order chromatin structure and genome stability, but how H1 functions in these processes is elusive. Here, we report that acetylation of lysine 85 (K85) within the H1 globular domain is a critical post-translational modification that regulates chromatin organization. H1K85 is dynamically acetylated by the acetyltransferase PCAF in response to DNA damage, and this effect is counterbalanced by the histone deacetylase HDAC1. Notably, an acetylation-mimic mutation of H1K85 (H1K85Q) alters H1 binding to the nucleosome and leads to condensed chromatin as a result of increased H1 binding to core histones. In addition, H1K85 acetylation promotes heterochromatin protein 1 (HP1) recruitment to facilitate chromatin compaction. Consequently, H1K85 mutation leads to genomic instability and decreased cell survival upon DNA damage. Together, our data suggest a novel model whereby H1K85 acetylation regulates chromatin structure and preserves chromosome integrity upon DNA damage.
Figures
References
-
- Fan Y., Nikitina T., Zhao J., Fleury T.J., Bhattacharyya R., Bouhassira E.E., Stein A., Woodcock C.L., Skoultchi A.I.. Histone H1 depletion in mammals alters global chromatin structure but causes specific changes in gene regulation. Cell. 2005; 123:1199–1212. - PubMed
-
- Song F., Chen P., Sun D., Wang M., Dong L., Liang D., Xu R.M., Zhu P., Li G.. Cryo-EM study of the chromatin fiber reveals a double helix twisted by tetranucleosomal units. Science. 2014; 344:376–380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
