

The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress
- PMID: 29983246
- PMCID: PMC6185997
- DOI: 10.1016/j.cmet.2018.06.008
The Mitochondrial-Encoded Peptide MOTS-c Translocates to the Nucleus to Regulate Nuclear Gene Expression in Response to Metabolic Stress
Abstract
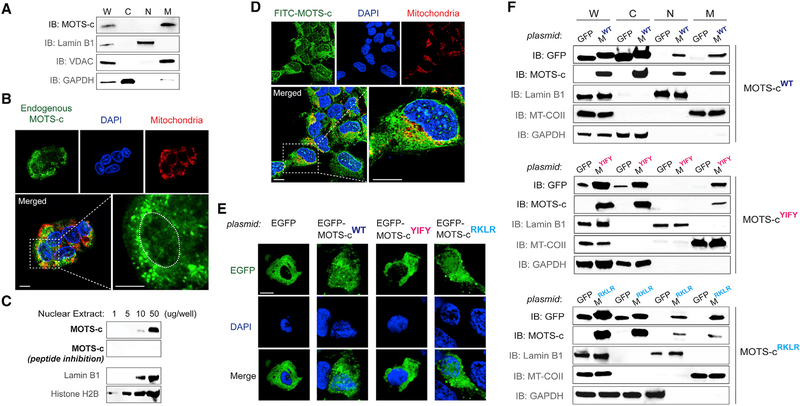
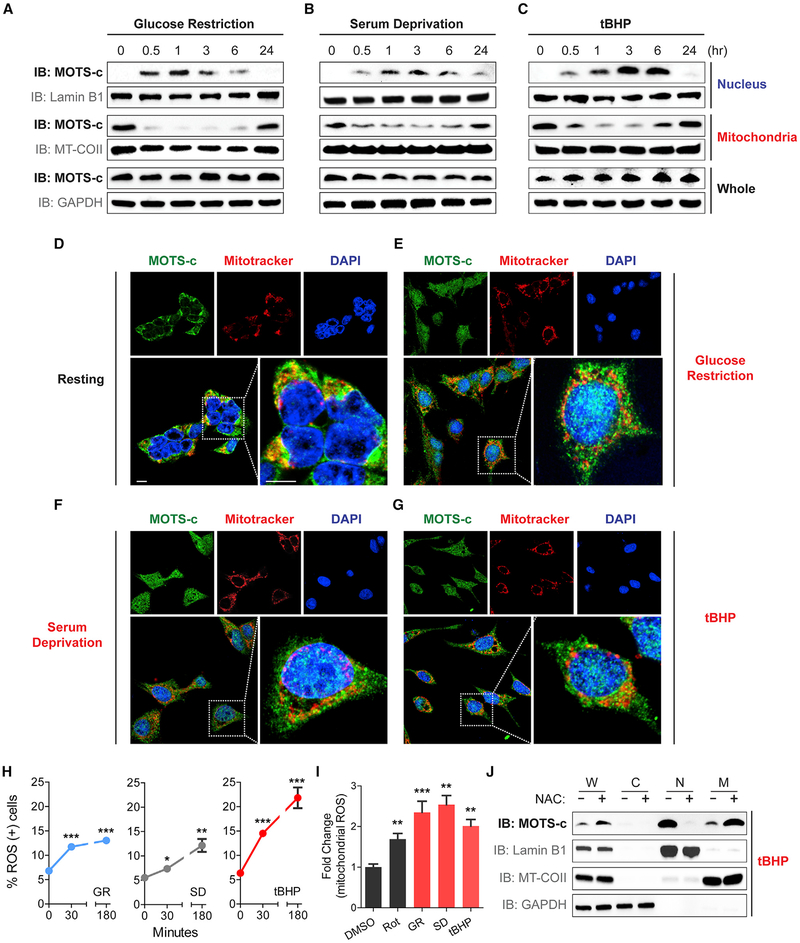
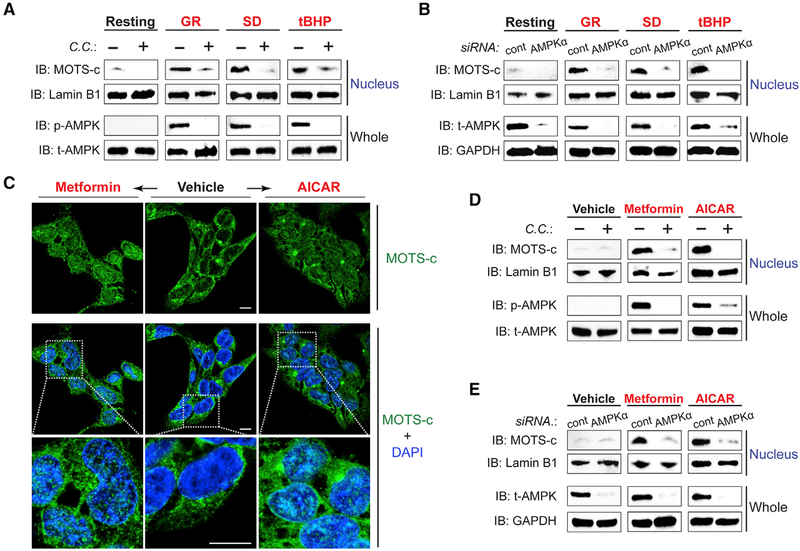
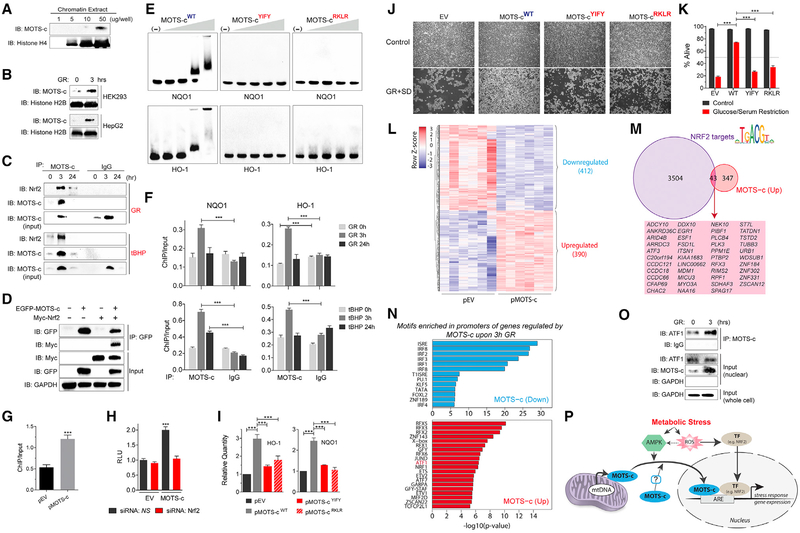
Cellular homeostasis is coordinated through communication between mitochondria and the nucleus, organelles that each possess their own genomes. Whereas the mitochondrial genome is regulated by factors encoded in the nucleus, the nuclear genome is currently not known to be actively controlled by factors encoded in the mitochondrial DNA. Here, we show that MOTS-c, a peptide encoded in the mitochondrial genome, translocates to the nucleus and regulates nuclear gene expression following metabolic stress in a 5'-adenosine monophosphate-activated protein kinase (AMPK)-dependent manner. In the nucleus, MOTS-c regulated a broad range of genes in response to glucose restriction, including those with antioxidant response elements (ARE), and interacted with ARE-regulating stress-responsive transcription factors, such as nuclear factor erythroid 2-related factor 2 (NFE2L2/NRF2). Our findings indicate that the mitochondrial and nuclear genomes co-evolved to independently encode for factors to cross-regulate each other, suggesting that mitonuclear communication is genetically integrated.
Keywords: MOTS-c; homeostasis; metabolic stress; mitochondria; mitochondrial DNA; mitochondrial-derived peptide (MDP); mitonuclear communication; peptide; short open reading frames (sORF); stress response.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
C.L. is a consultant for and a shareholder of CohBar, Inc. The remaining authors declare no competing interests.
Figures
Comment in
-
A Mitochondrial-Derived Peptide Exercises the Nuclear Option.Cell Metab. 2018 Sep 4;28(3):330-331. doi: 10.1016/j.cmet.2018.08.017. Cell Metab. 2018. PMID: 30184481
References
-
- Bray NL, Pimentel H, Melsted P, and Pachter L (2016). Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol 34, 525–527. - PubMed
-
- Brunmair B, Staniek K, Gras F, Scharf N, Althaym A, Clara R, Roden M, Gnaiger E, Nohl H, Waldhäusl W, and Fürnsinn C (2004). Thiazolidinediones, like metformin, inhibit respiratory complex I: a common mechanism contributing to their antidiabetic actions? Diabetes 53, 1052–1059. - PubMed
-
- Chandel NS (2015). Evolution of mitochondria as signaling organelles. Cell Metab. 22, 204–206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
