

A Quantitative Chemoproteomic Platform to Monitor Selenocysteine Reactivity within a Complex Proteome
- PMID: 29983274
- PMCID: PMC6510024
- DOI: 10.1016/j.chembiol.2018.05.017
A Quantitative Chemoproteomic Platform to Monitor Selenocysteine Reactivity within a Complex Proteome
Abstract
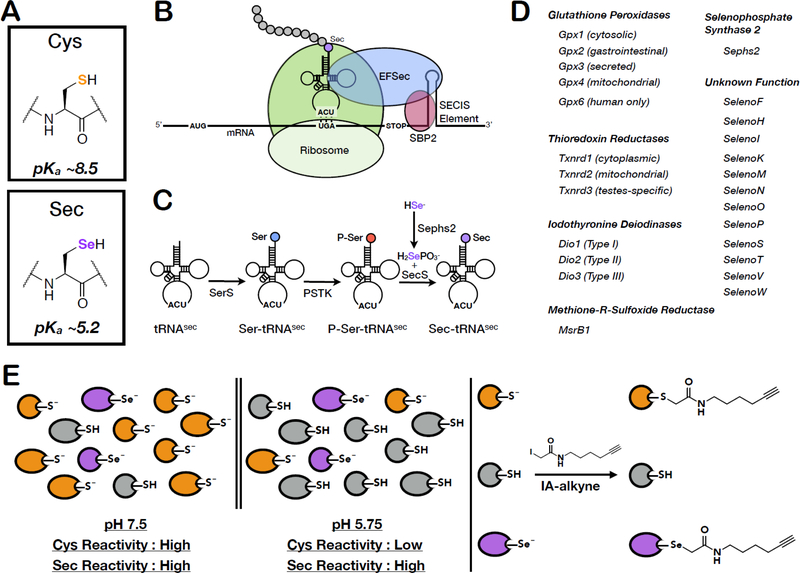
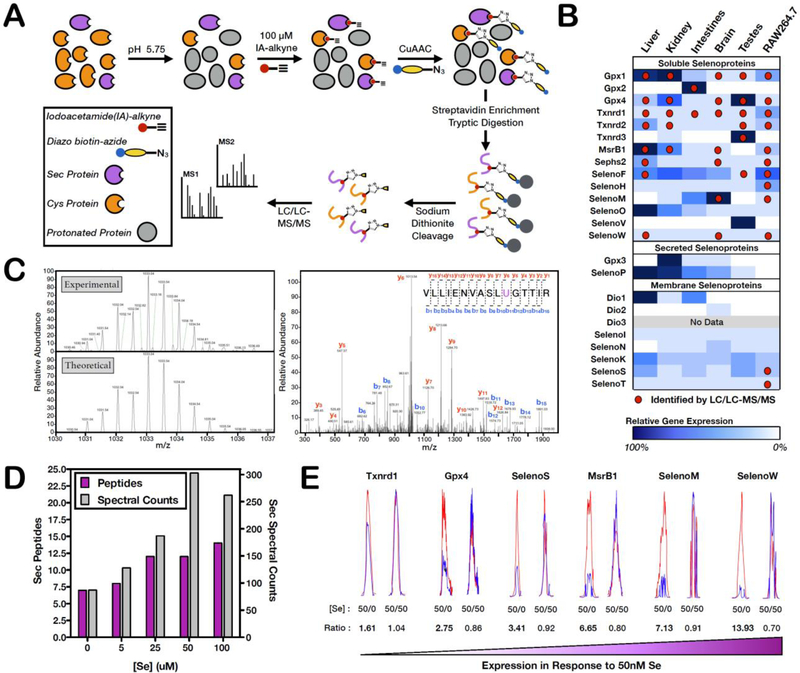
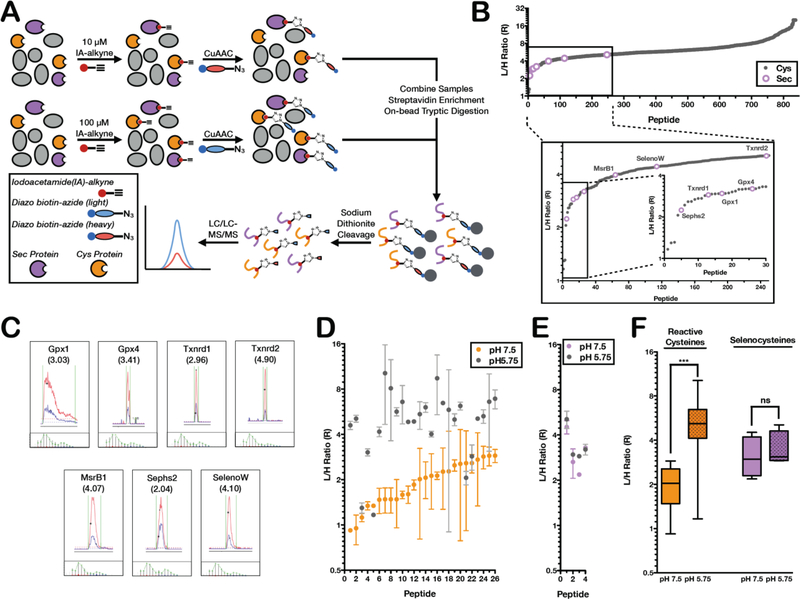
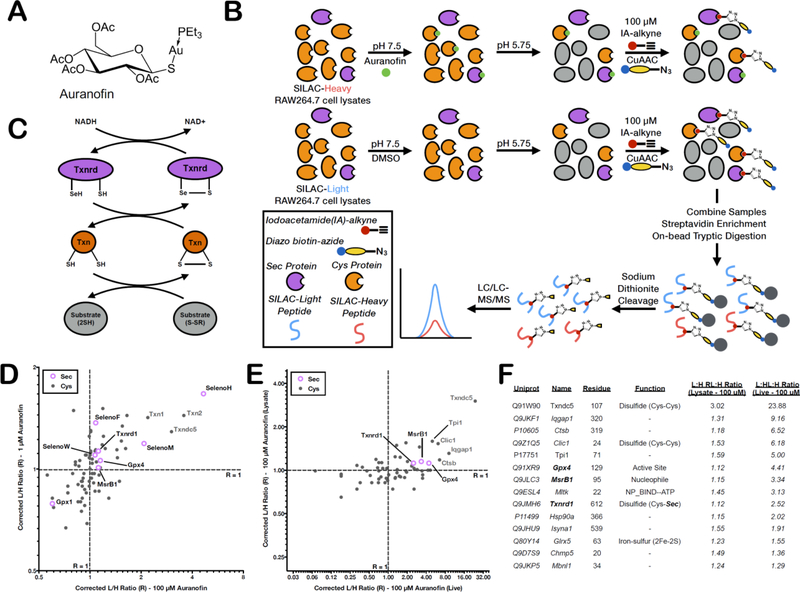
Mammalian selenocysteine (Sec)-containing proteins, selenoproteins, are important to (patho)physiological processes, including redox homeostasis. Sec residues have been recalcitrant to mass spectrometry-based chemoproteomic methods that enrich for reactive cysteine (Cys) residues with electrophilic chemical probes, despite confirmed reactivity of Sec with these electrophiles. Highly abundant Cys peptides likely suppress low-abundant Sec peptides. By exploiting the decreased pKa of Sec relative to Cys, we have developed a chemoproteomic platform that relies on low pH (pH 5.75) electrophile labeling, reducing Cys reactivity and enhancing identification of Sec-containing peptides across mouse tissues and cell lines. The utility of this Sec-profiling platform is underscored by evaluation of the selectivity of auranofin, an inhibitor of the selenoprotein, thioredoxin reductase, against both reactive Cys- and Sec-containing proteins. Platform limitations pertain to the non-physiological low-pH conditions that could perturb protein structure and function. Future work necessitates the discovery of Sec-selective electrophiles that function at physiological pH.
Keywords: chemoproteomics; cysteine; isoTOP-ABPP; selenocysteine; selenoprotein.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
References
-
- Al-Taie OH, Uceyler N, Eubner U, Jakob F, Mork H, Scheurlen M, Brigelius-Flohe R, Schottker K, Abel J, Thalheimer A, et al. (2009). Expression Profiling and Genetic Alterations of the Selenoproteins GI-GPx and SePP in Colorectal Carcinogenesis. Nutr. Cancer 48, 6–14. - PubMed
-
- Apostolou S, Klein JO, Mitsuuchi Y, and Shetler JN (2004). Growth inhibition and induction of apoptosis in mesothelioma cells by selenium and dependence on selenoprotein SEP15 genotype. Oncogene 23, 5032–5040. - PubMed
-
- Arnér ESJ (2010). Selenoproteins—What unique properties can arise with selenocysteine in place of cysteine? Exp. Cell Res 316, 1296–1303. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
