

AMPA receptor trafficking and its role in heterosynaptic plasticity
- PMID: 29985438
- PMCID: PMC6037747
- DOI: 10.1038/s41598-018-28581-w
AMPA receptor trafficking and its role in heterosynaptic plasticity
Abstract
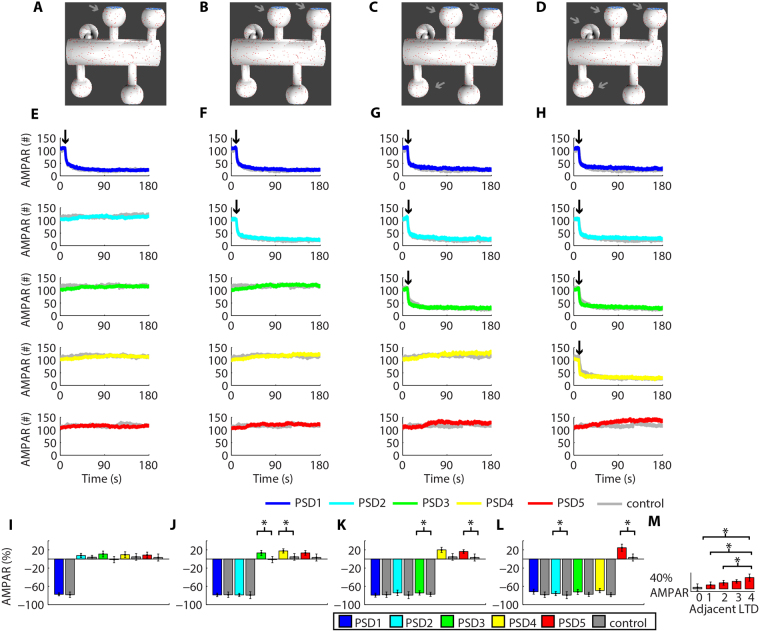
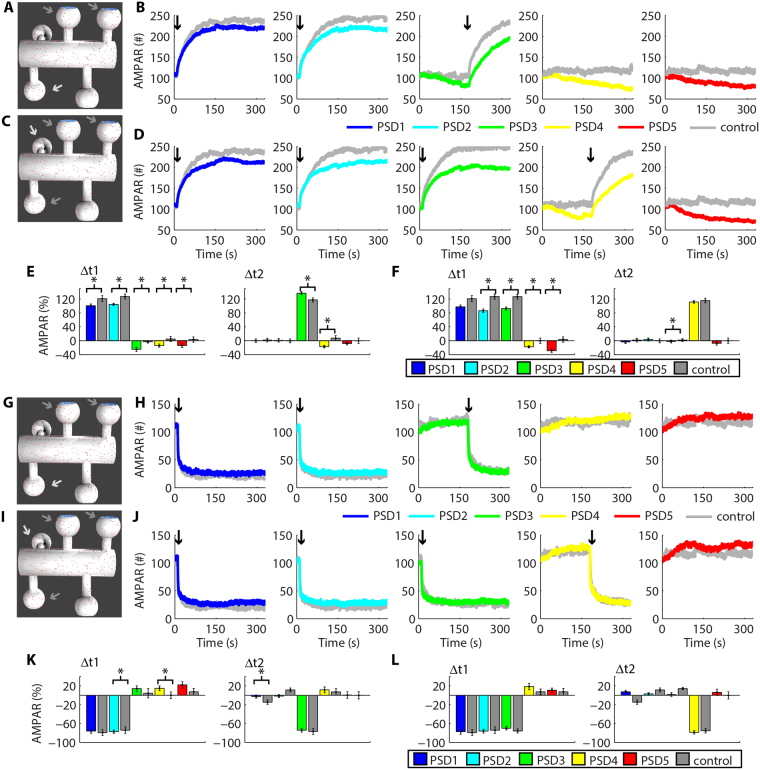
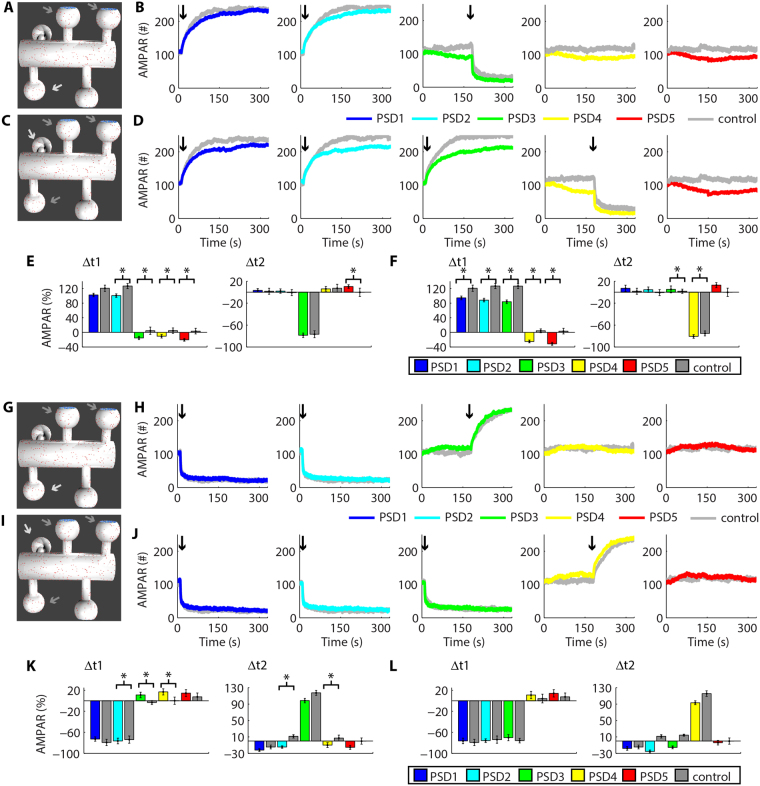
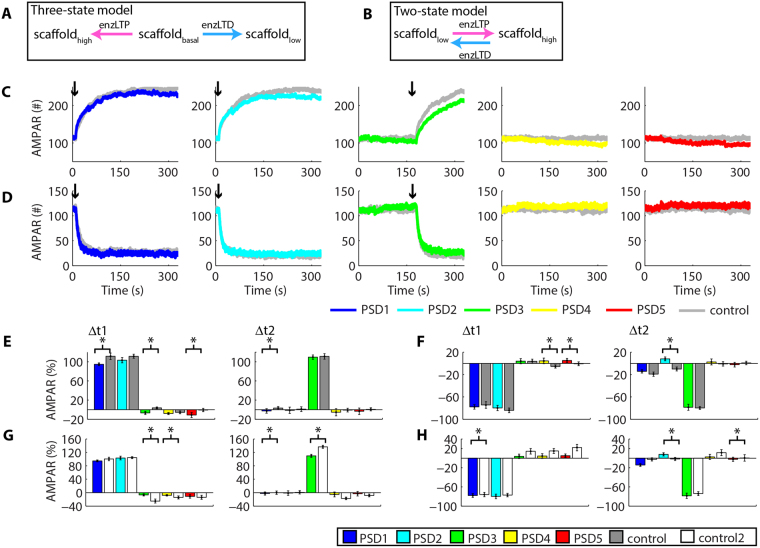
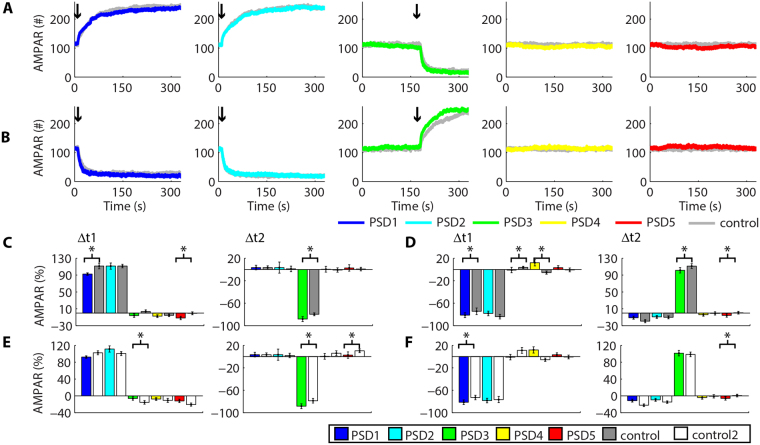
Historically, long-term potentiation (LTP) and long-term depression (LTD), the best-characterized forms of long-term synaptic plasticity, are viewed as experience-dependent and input-specific processes. However, cumulative experimental and theoretical data have demonstrated that LTP and LTD can promote compensatory alterations in non-stimulated synapses. In this work, we have developed a computational model of a tridimensional spiny dendritic segment to investigate the role of AMPA receptor (AMPAR) trafficking during synaptic plasticity at specific synapses and its consequences for the populations of AMPAR at nearby synapses. Our results demonstrated that the mechanisms of AMPAR trafficking involved with LTP and LTD can promote heterosynaptic plasticity at non-stimulated synapses. These alterations are compensatory and arise from molecular competition. Moreover, the heterosynaptic changes observed in our model can modulate further activity-driven inductions of synaptic plasticity.
Conflict of interest statement
The authors declare no competing interests.
Figures
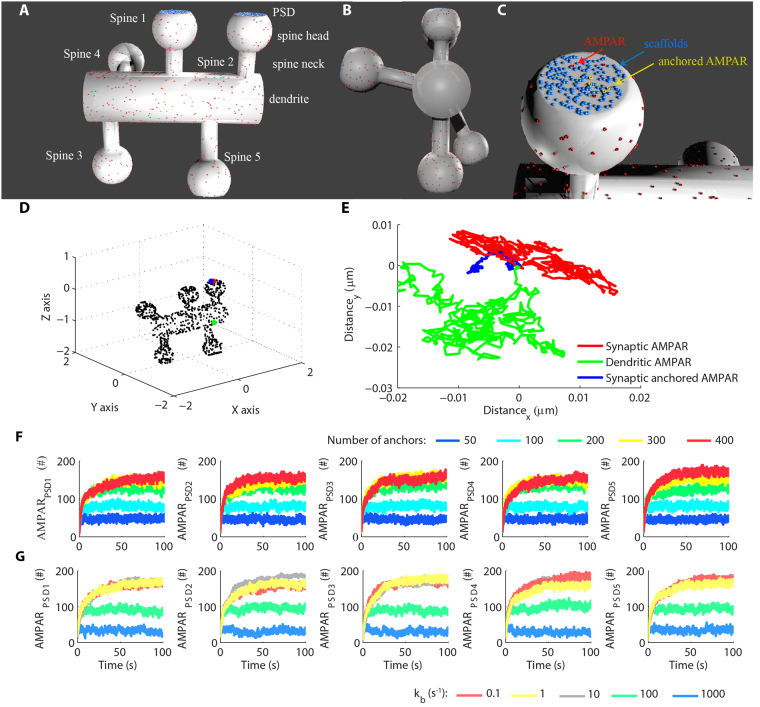
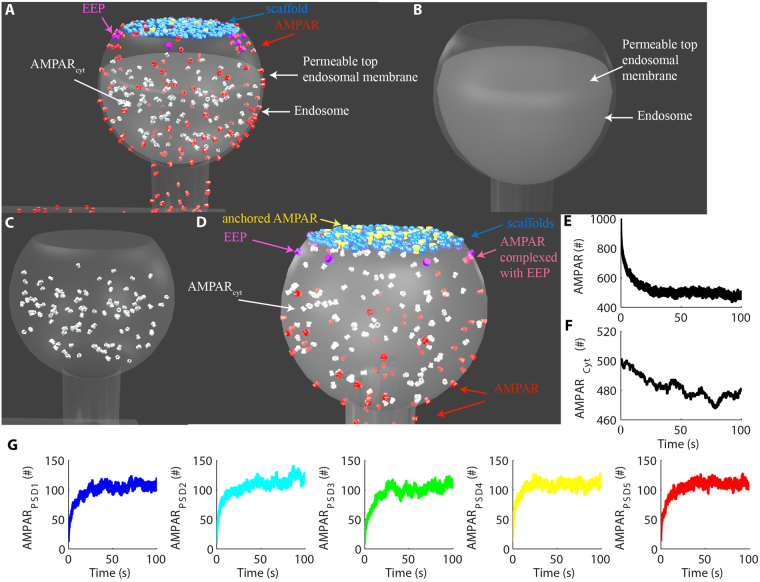
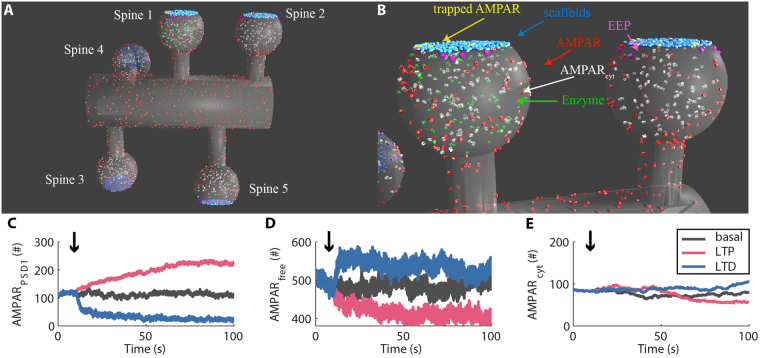
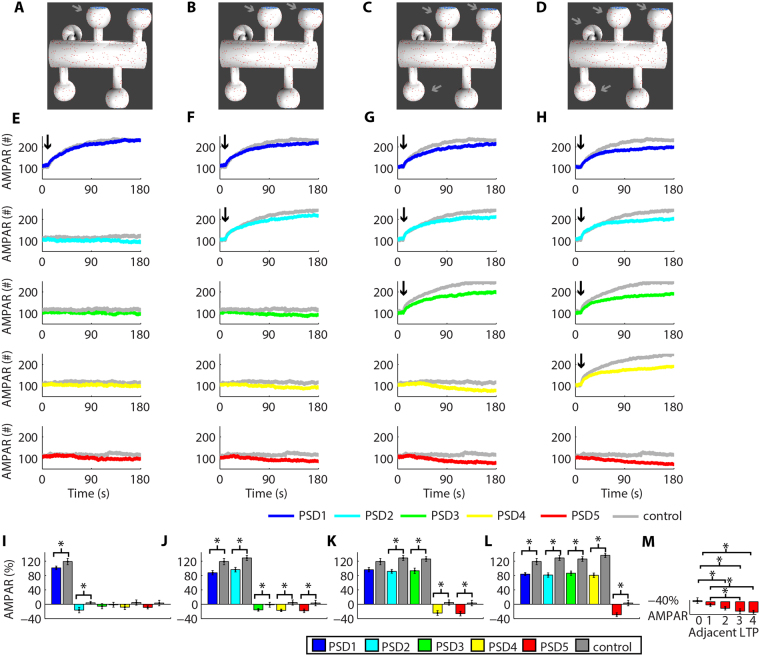
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
