

GROWTH-REGULATING FACTOR 9 negatively regulates arabidopsis leaf growth by controlling ORG3 and restricting cell proliferation in leaf primordia
- PMID: 29985961
- PMCID: PMC6053248
- DOI: 10.1371/journal.pgen.1007484
GROWTH-REGULATING FACTOR 9 negatively regulates arabidopsis leaf growth by controlling ORG3 and restricting cell proliferation in leaf primordia
Abstract
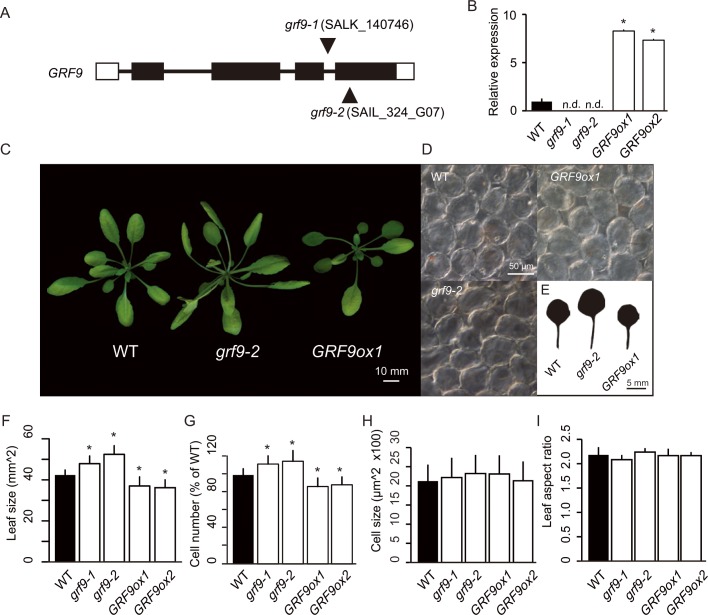
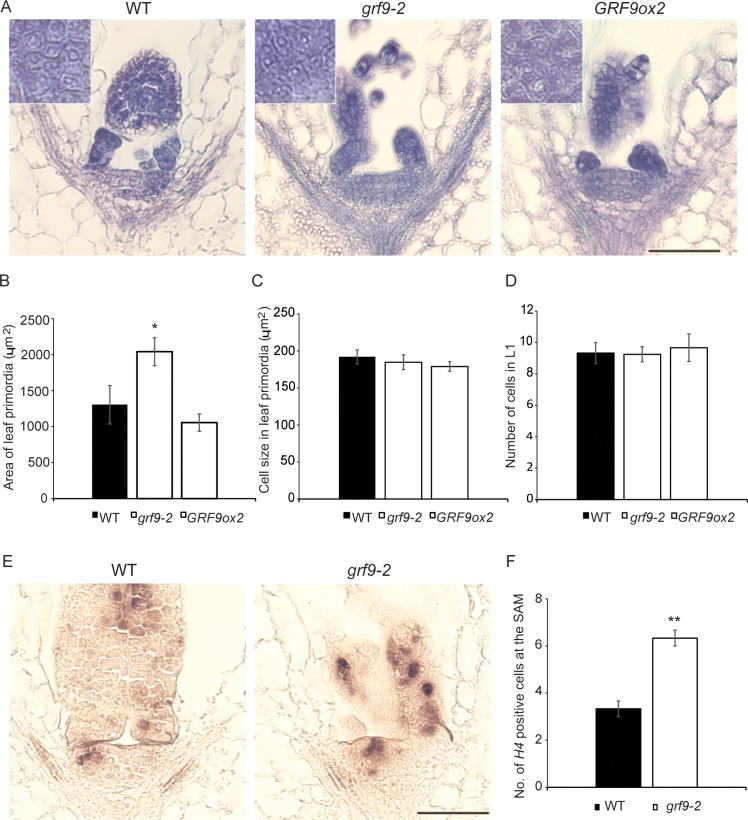
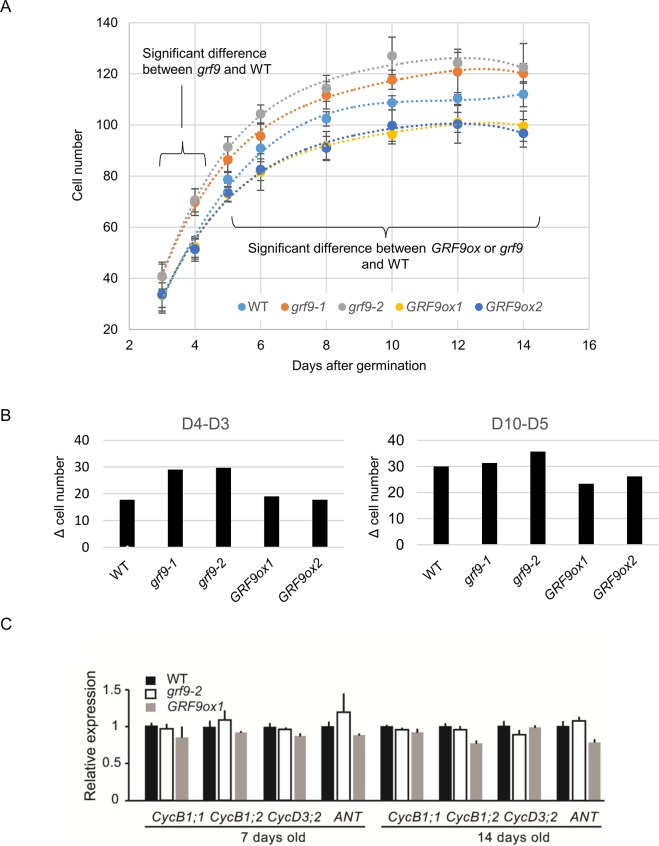
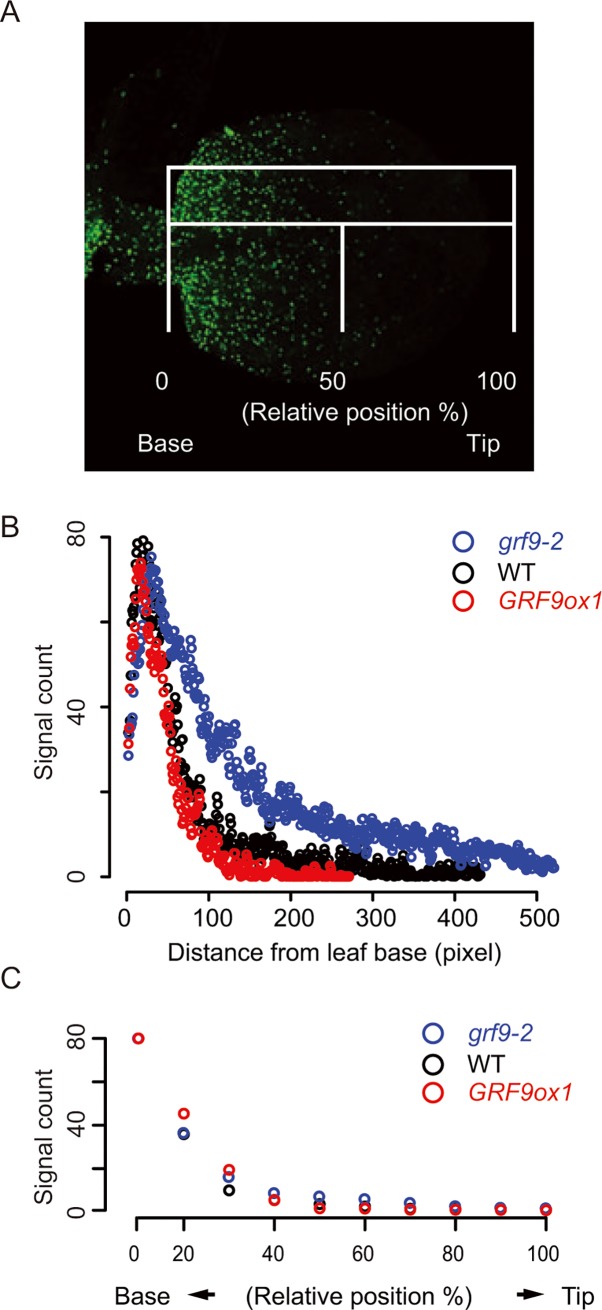
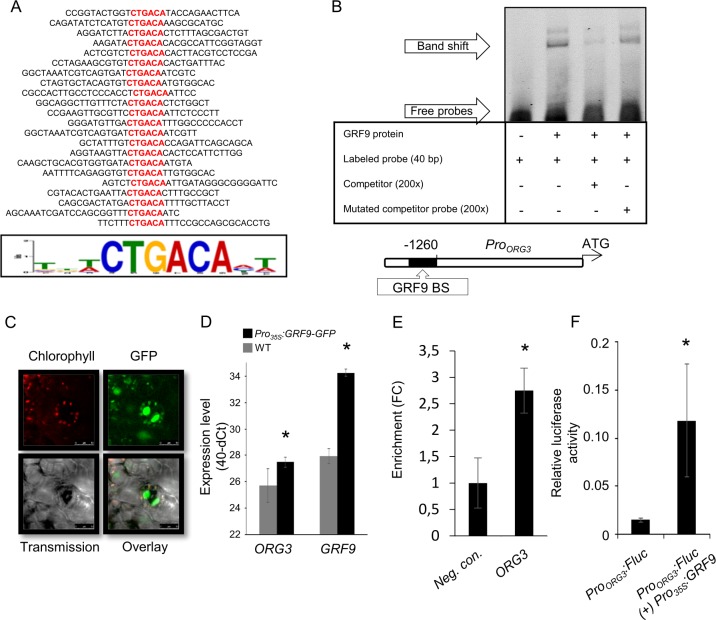
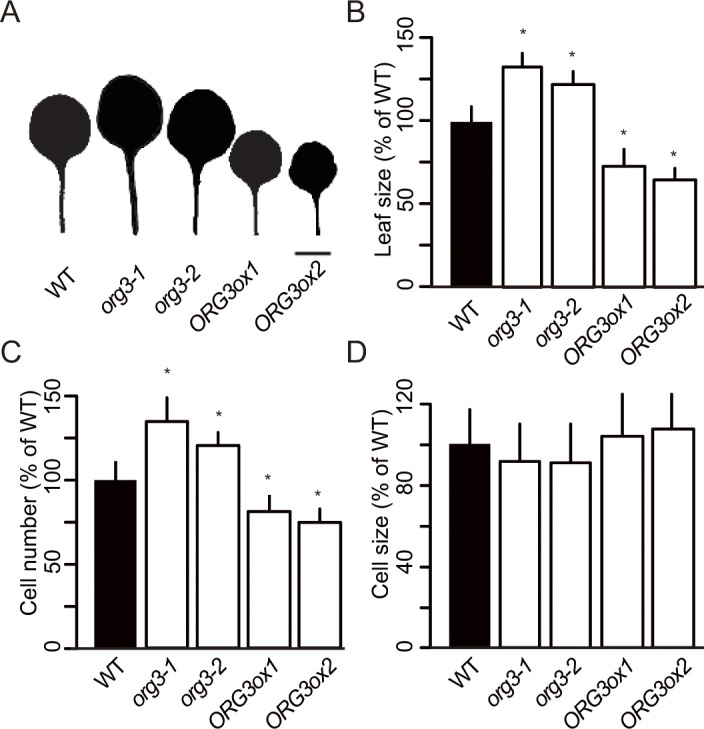
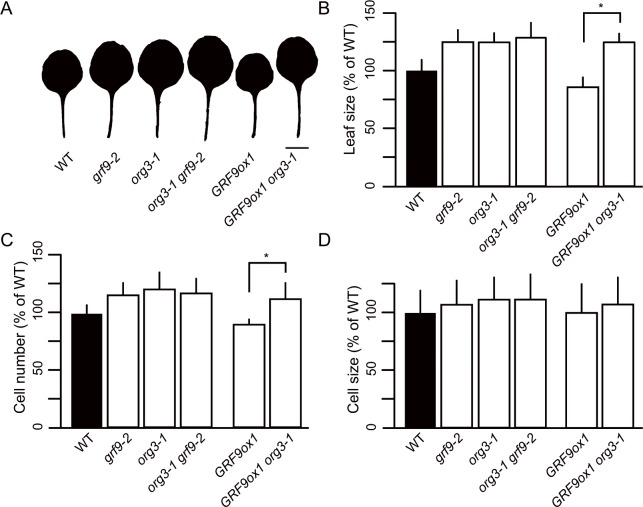
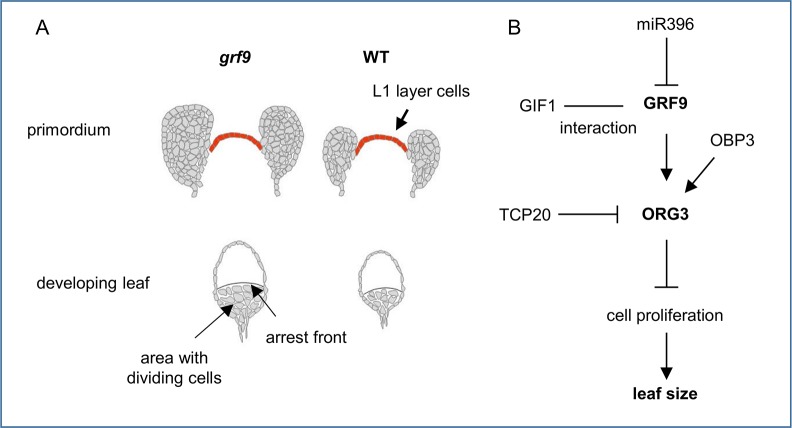
Leaf growth is a complex process that involves the action of diverse transcription factors (TFs) and their downstream gene regulatory networks. In this study, we focus on the functional characterization of the Arabidopsis thaliana TF GROWTH-REGULATING FACTOR9 (GRF9) and demonstrate that it exerts its negative effect on leaf growth by activating expression of the bZIP TF OBP3-RESPONSIVE GENE 3 (ORG3). While grf9 knockout mutants produce bigger incipient leaf primordia at the shoot apex, rosette leaves and petals than the wild type, the sizes of those organs are reduced in plants overexpressing GRF9 (GRF9ox). Cell measurements demonstrate that changes in leaf size result from alterations in cell numbers rather than cell sizes. Kinematic analysis and 5-ethynyl-2'-deoxyuridine (EdU) incorporation assay revealed that GRF9 restricts cell proliferation in the early developing leaf. Performing in vitro binding site selection, we identified the 6-base motif 5'-CTGACA-3' as the core binding site of GRF9. By global transcriptome profiling, electrophoretic mobility shift assay (EMSA) and chromatin immunoprecipitation (ChIP) we identified ORG3 as a direct downstream, and positively regulated target of GRF9. Genetic analysis of grf9 org3 and GRF9ox org3 double mutants reveals that both transcription factors act in a regulatory cascade to control the final leaf dimensions by restricting cell number in the developing leaf.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
