

MetClo: methylase-assisted hierarchical DNA assembly using a single type IIS restriction enzyme
- PMID: 29986052
- PMCID: PMC6212791
- DOI: 10.1093/nar/gky596
MetClo: methylase-assisted hierarchical DNA assembly using a single type IIS restriction enzyme
Abstract
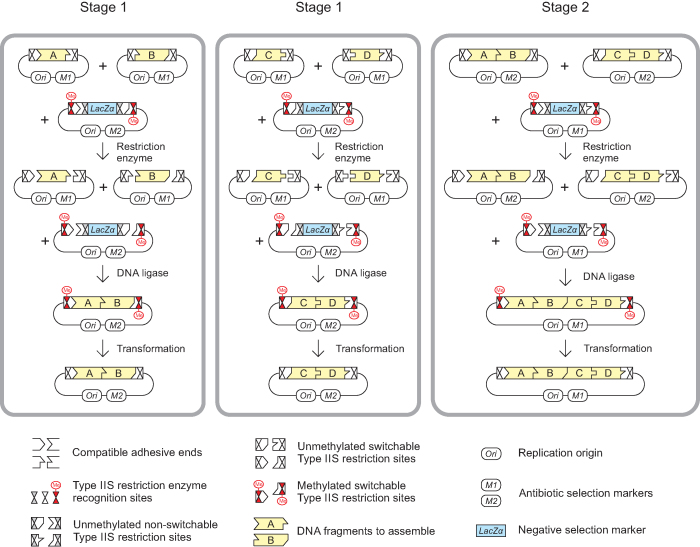
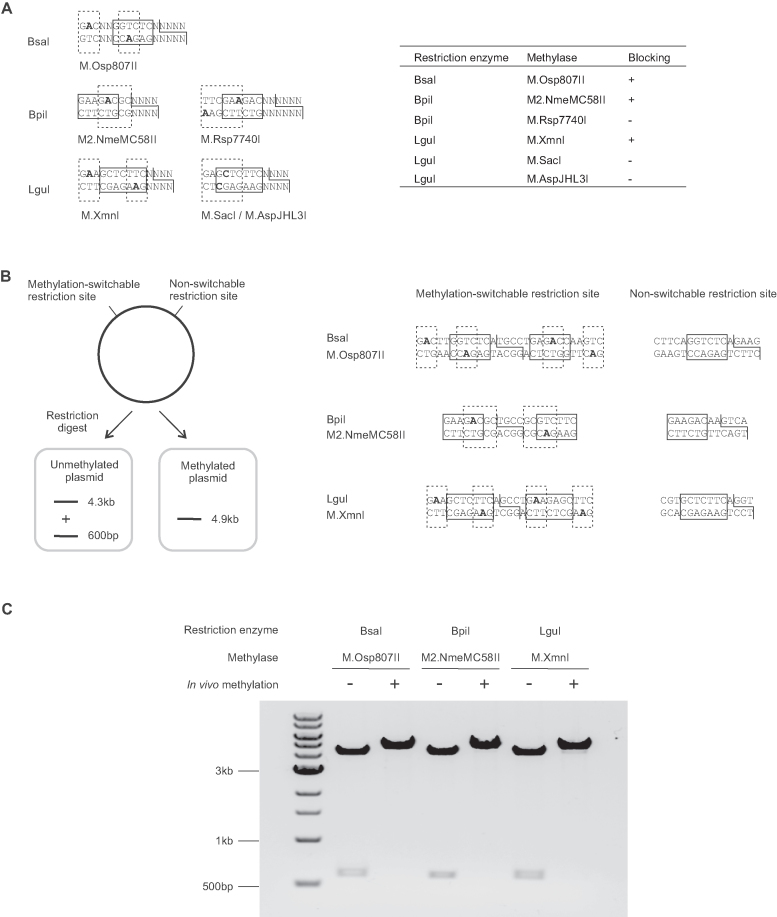
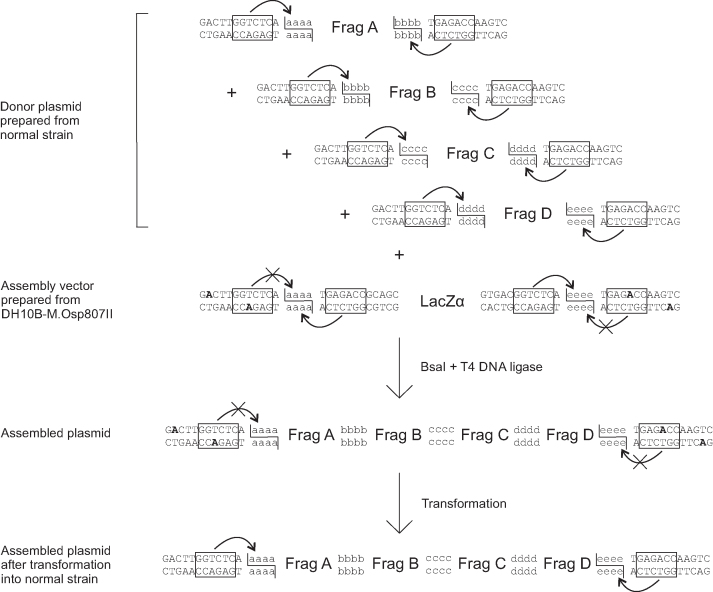
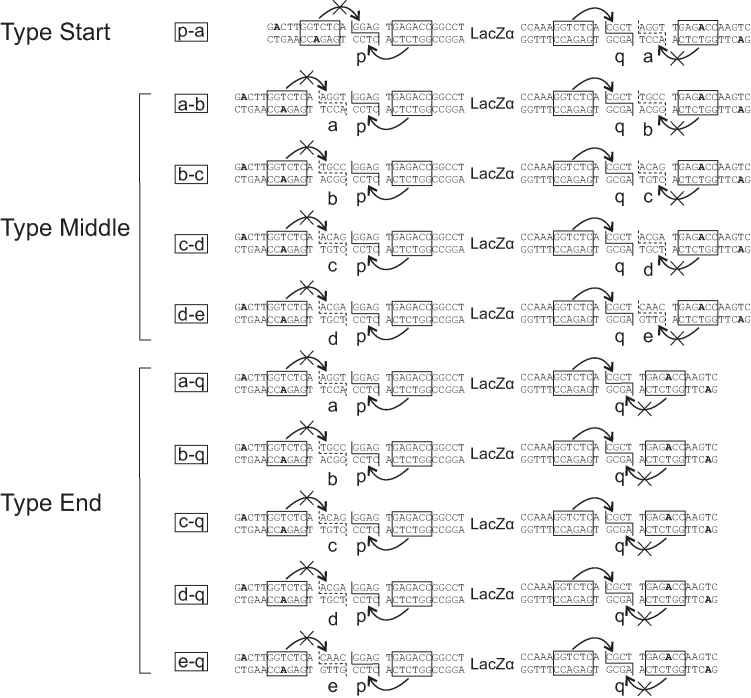
Efficient DNA assembly is of great value in biological research and biotechnology. Type IIS restriction enzyme-based assembly systems allow assembly of multiple DNA fragments in a one-pot reaction. However, large DNA fragments can only be assembled by alternating use of two or more type IIS restriction enzymes in a multi-step approach. Here, we present MetClo, a DNA assembly method that uses only a single type IIS restriction enzyme for hierarchical DNA assembly. The method is based on in vivo methylation-mediated on/off switching of type IIS restriction enzyme recognition sites that overlap with site-specific methylase recognition sequences. We have developed practical MetClo systems for the type IIS enzymes BsaI, BpiI and LguI, and demonstrated hierarchical assembly of large DNA fragments up to 218 kb. The MetClo approach substantially reduces the need to remove internal restriction sites from components to be assembled. The use of a single type IIS enzyme throughout the different stages of DNA assembly allows novel and powerful design schemes for rapid large-scale hierarchical DNA assembly. The BsaI-based MetClo system is backward-compatible with component libraries of most of the existing type IIS restriction enzyme-based assembly systems, and has potential to become a standard for modular DNA assembly.
Figures
References
-
- Casini A., Storch M., Baldwin G.S., Ellis T.. Bricks and blueprints: methods and standards for DNA assembly. Nat. Rev. Mol. Cell Biol. 2015; 16:568–576. - PubMed
-
- Juhas M., Ajioka J.W.. High molecular weight DNA assembly in vivo for synthetic biology applications. Crit. Rev. Biotechnol. 2017; 37:277–286. - PubMed
-
- Gibson D.G., Young L., Chuang R.Y., Venter J.C., Hutchison C.A. 3rd, Smith H.O.. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods. 2009; 6:343–345. - PubMed
-
- Itaya M., Fujita K., Kuroki A., Tsuge K.. Bottom-up genome assembly using the Bacillus subtilis genome vector. Nat. Methods. 2008; 5:41–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
