

Highly focused transcriptional response of Anopheles coluzzii to O'nyong nyong arbovirus during the primary midgut infection
- PMID: 29986645
- PMCID: PMC6038350
- DOI: 10.1186/s12864-018-4918-0
Highly focused transcriptional response of Anopheles coluzzii to O'nyong nyong arbovirus during the primary midgut infection
Abstract
Background: Anopheles mosquitoes are efficient vectors of human malaria, but it is unknown why they do not transmit viruses as well as Aedes and Culex mosquitoes. The only arbovirus known to be consistently transmitted by Anopheles mosquitoes is O'nyong nyong virus (ONNV, genus Alphavirus, family Togaviridae). The interaction of Anopheles mosquitoes with RNA viruses has been relatively unexamined.
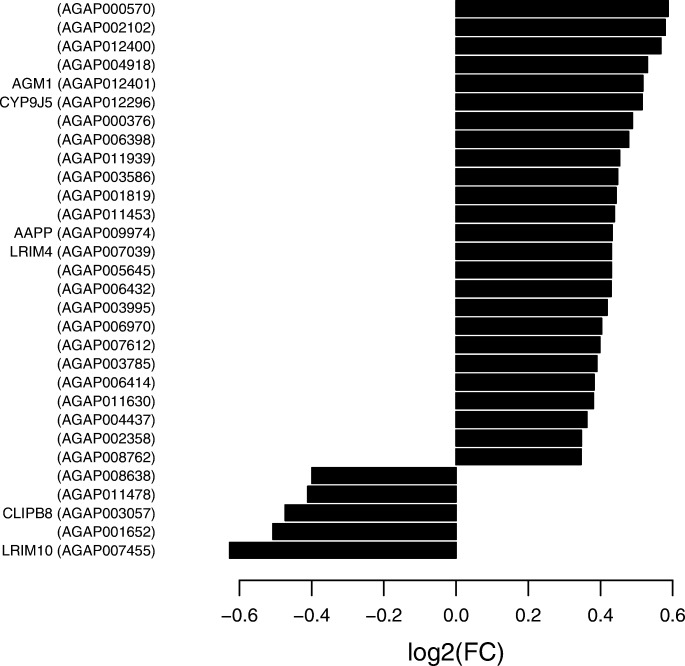
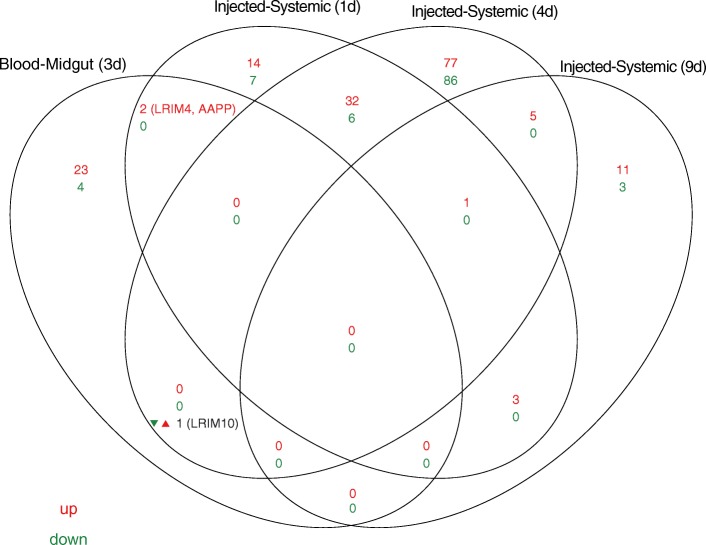
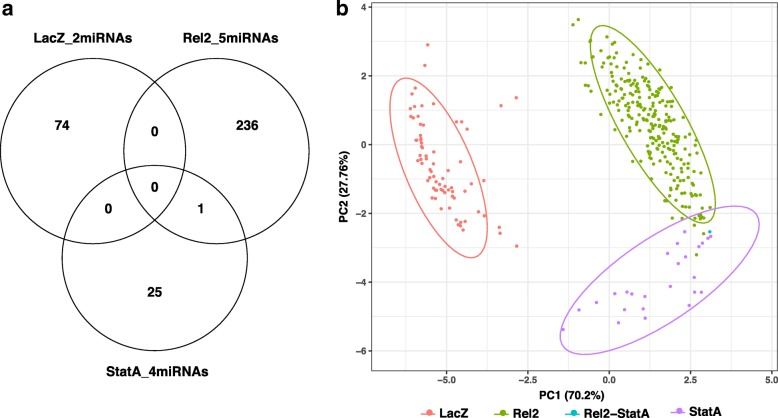
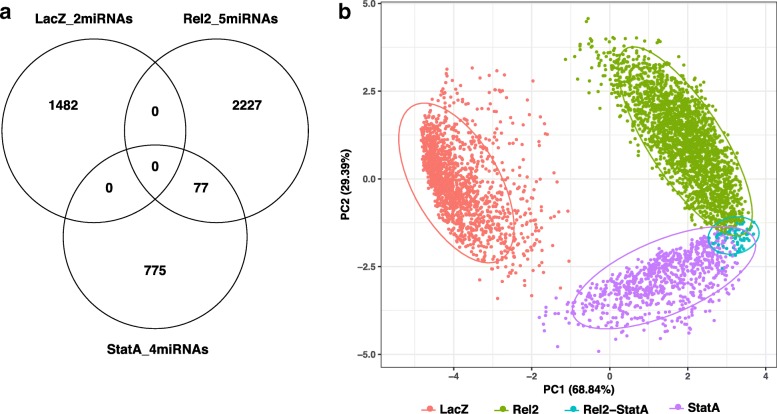
Results: We transcriptionally profiled the African malaria vector, Anopheles coluzzii, infected with ONNV. Mosquitoes were fed on an infectious bloodmeal and were analyzed by Illumina RNAseq at 3 days post-bloodmeal during the primary virus infection of the midgut epithelium, before systemic dissemination. Virus infection triggers transcriptional regulation of just 30 host candidate genes. Most of the regulated candidate genes are novel, without known function. Of the known genes, a significant cluster includes candidates with predicted involvement in carbohydrate metabolism. Two candidate genes encoding leucine-rich repeat immune (LRIM) factors point to possible involvement of immune protein complexes in the mosquito antiviral response. The primary ONNV infection by bloodmeal shares little transcriptional response in common with ONNV infection by intrathoracic injection, nor with midgut infection by the malaria parasites, Plasmodium falciparum or P. berghei. Profiling of A. coluzzii microRNA (miRNA) identified 118 known miRNAs and 182 potential novel miRNA candidates, with just one miRNA regulated by ONNV infection. This miRNA was not regulated by other previously reported treatments, and may be virus specific. Coexpression analysis of miRNA abundance and messenger RNA expression revealed discrete clusters of genes regulated by Imd and JAK/STAT, immune signaling pathways that are protective against ONNV in the primary infection.
Conclusions: ONNV infection of the A. coluzzii midgut triggers a remarkably limited gene regulation program of mostly novel candidate genes, which likely includes host genes deployed for antiviral defense, as well as genes manipulated by the virus to facilitate infection. Functional dissection of the ONNV-response candidate genes is expected to generate novel insight into the mechanisms of virus-vector interaction.
Keywords: Arbovirus; Host–pathogen interactions; Innate immunity; Insect immunity; Insect vectors; Malaria.
Conflict of interest statement
Ethics approval and consent to participate
There were no human subjects. The protocol for the ethical treatment of the animals used in this study was approved by the research animal ethics committee of the Institut Pasteur, “C2EA-89 CETEA Institut Pasteur” as protocol number B75–15-31. The Institut Pasteur ethics committee is authorized by the French Ministry of Higher Education and Research (MESR) under French law N° 2001–486, which is aligned with Directive 2010/63/EU of the European Commission on the protection of animals used for scientific purposes.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Atkins GJ. The pathogenesis of alphaviruses. ISRN Virology. 2013;2013:1–22. doi: 10.5402/2013/861912. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
