

Unveiling anti-oxidative and anti-inflammatory effects of docosahexaenoic acid and its lipid peroxidation product on lipopolysaccharide-stimulated BV-2 microglial cells
- PMID: 29986724
- PMCID: PMC6038194
- DOI: 10.1186/s12974-018-1232-3
Unveiling anti-oxidative and anti-inflammatory effects of docosahexaenoic acid and its lipid peroxidation product on lipopolysaccharide-stimulated BV-2 microglial cells
Abstract
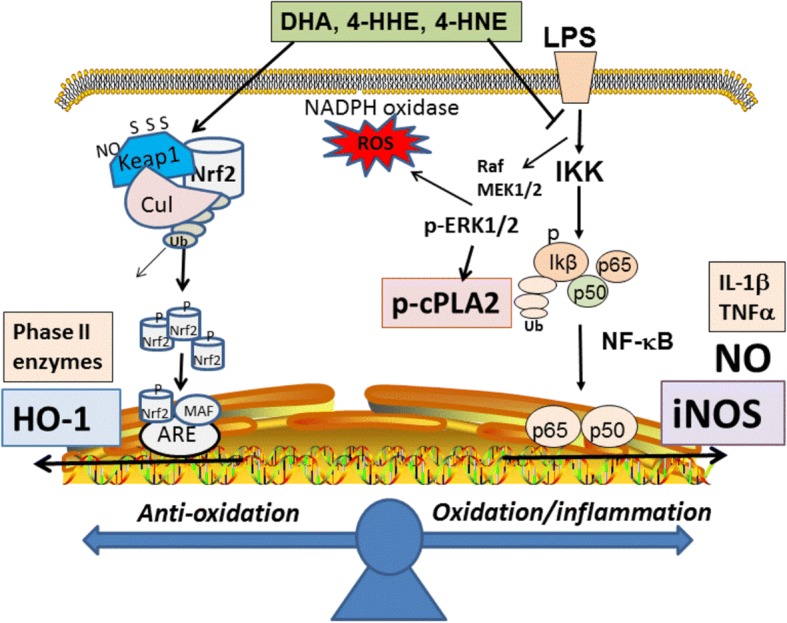
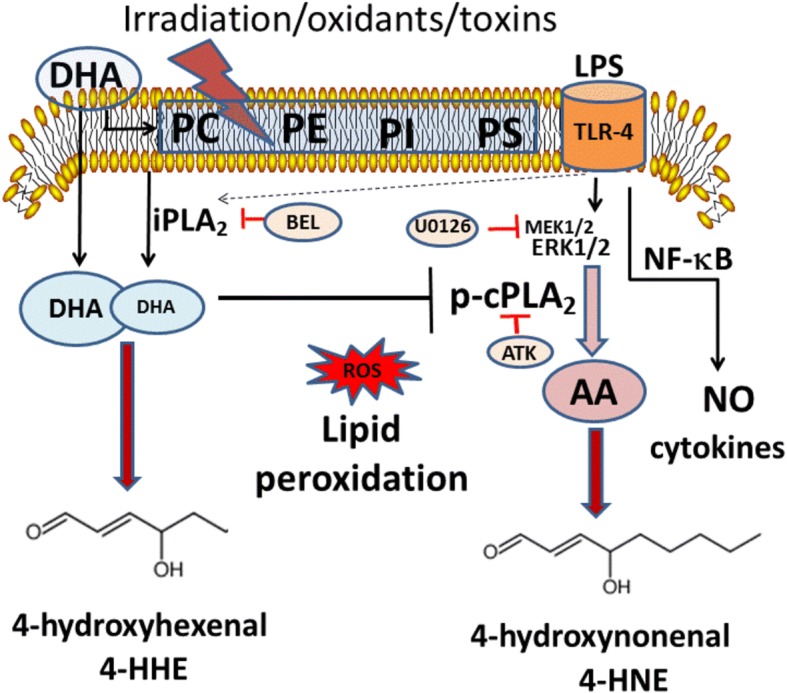
Background: Phospholipids in the central nervous system are enriched in n-3 and n-6 polyunsaturated fatty acids (PUFA), especially docosahexaenoic acid (DHA) and arachidonic acid (ARA). These PUFA can undergo enzymatic reactions to produce lipid mediators, as well as reaction with oxygen free radicals to produce 4-hydroxyhexenal (4-HHE) from DHA and 4-hydroxynonenal (4-HNE) from ARA. Recent studies demonstrated pleiotropic properties of these peroxidation products through interaction with oxidative and anti-oxidant response pathways. In this study, BV-2 microglial cells were used to investigate ability for DHA, 4-HHE, and 4-HNE to stimulate the anti-oxidant stress responses involving the nuclear factor erythroid-2-related factor 2 (Nrf2) pathway and synthesis of heme oxygenase (HO-1), as well as to mitigate lipopolysaccharide (LPS)-induced nitric oxide (NO), reactive oxygen species (ROS), and cytosolic phospholipase A2 (cPLA2). In addition, LC-MS/MS analysis was carried out to examine effects of exogenous DHA and LPS stimulation on endogenous 4-HHE and 4-HNE levels in BV-2 microglial cells.
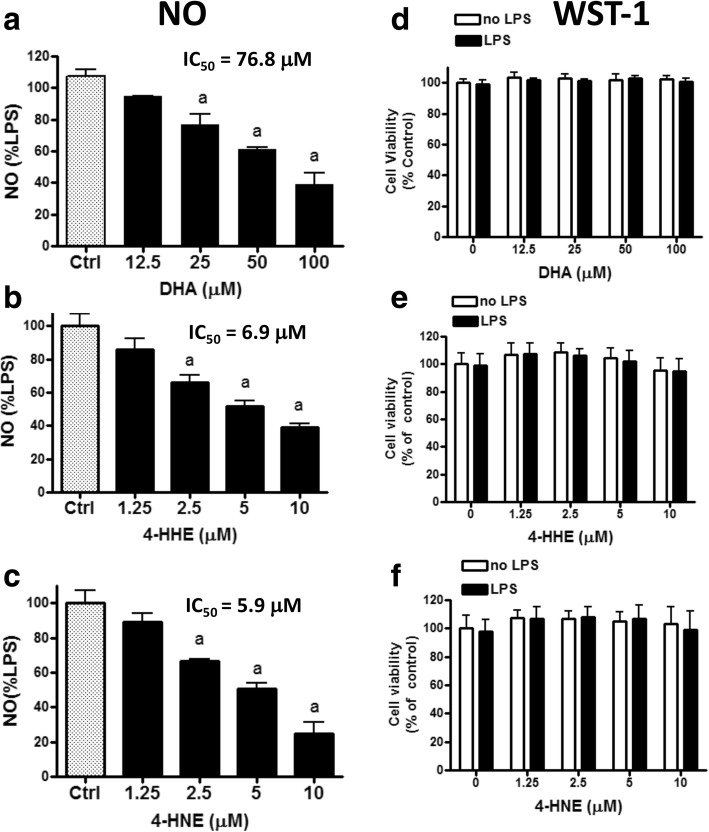
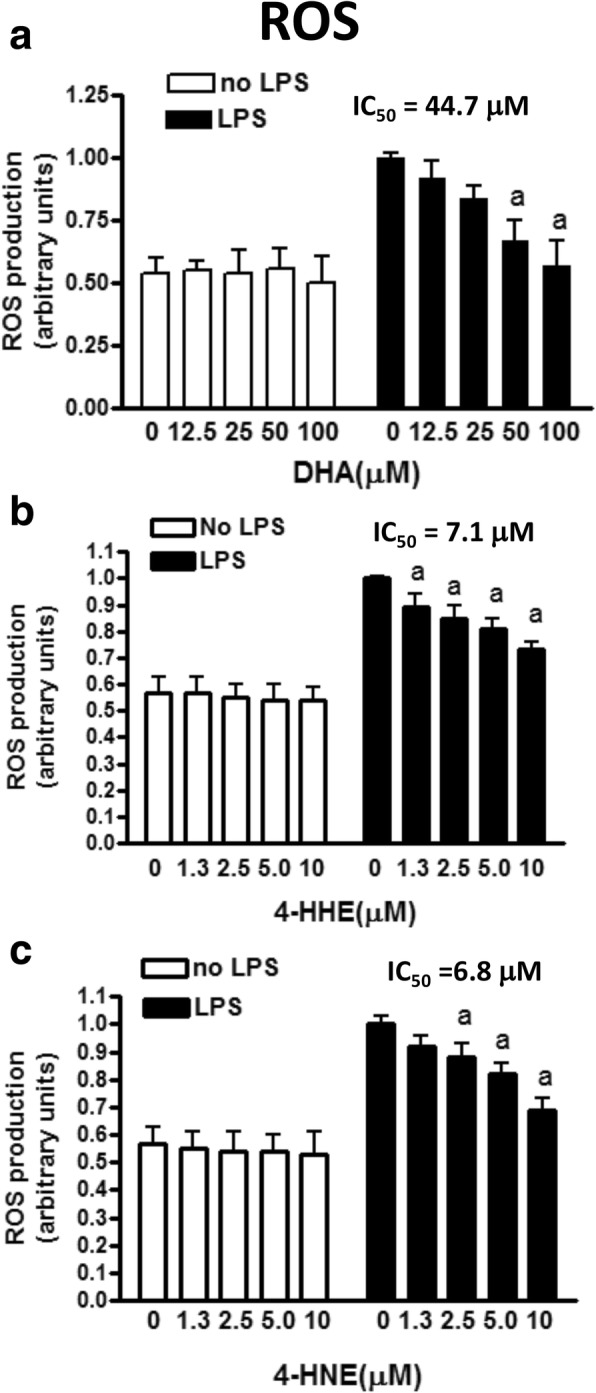
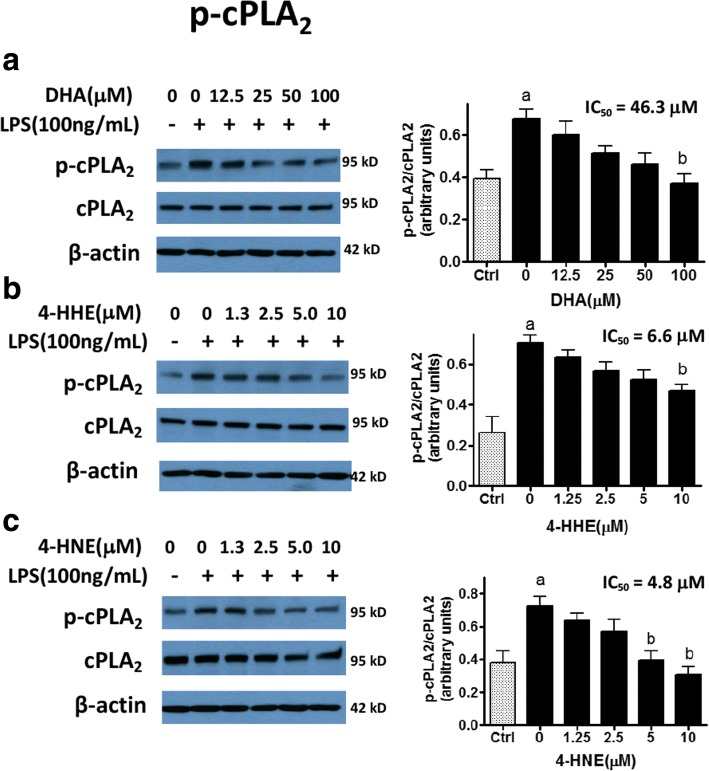
Methods: Effects of DHA, 4-HHE, and 4-HNE on LPS-induced NO production was determined using the Griess reagent. LPS-induced ROS production was measured using CM-H2DCFDA. Western blots were used to analyze expression of p-cPLA2, Nrf2, and HO-1. Cell viability and cytotoxicity were measured using the WST-1 assay, and cell protein concentrations were measured using the BCA protein assay kit. An ultra-high-performance liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis was used to determine levels of free 4-HHE and 4-HNE in cells.
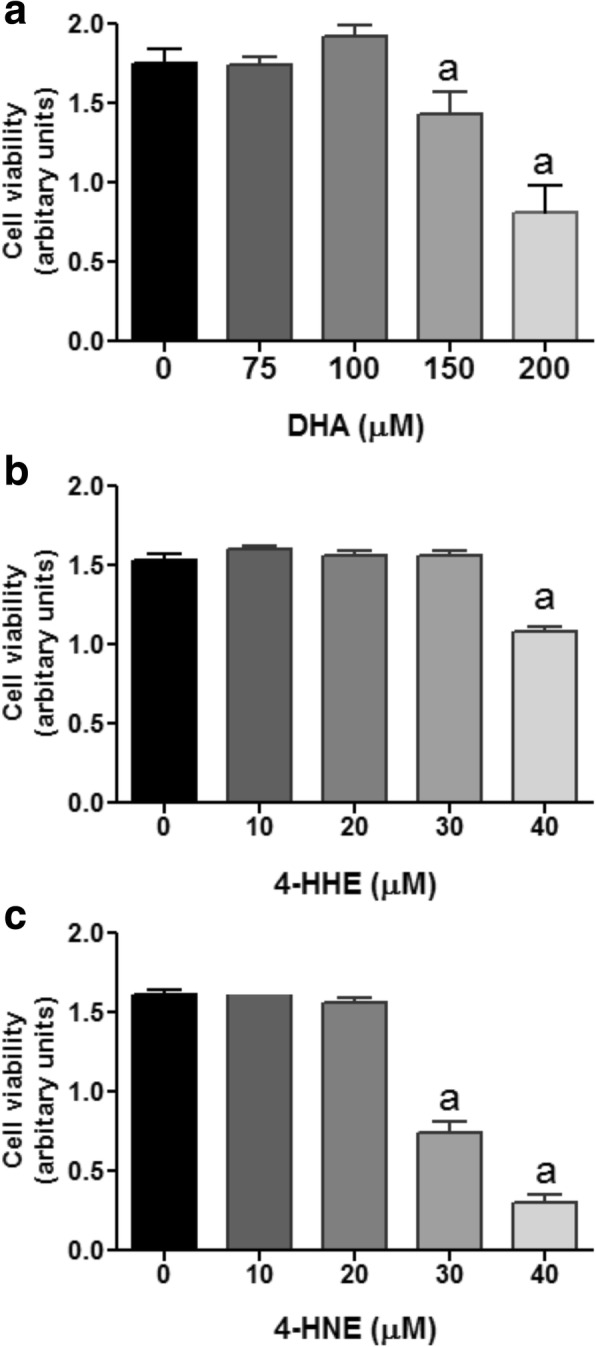
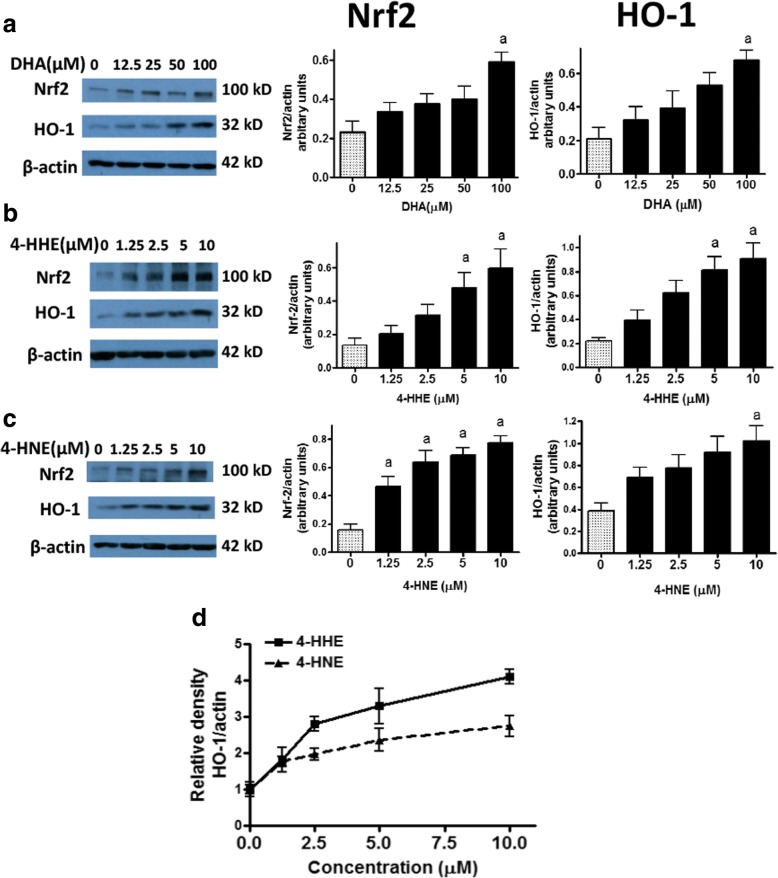
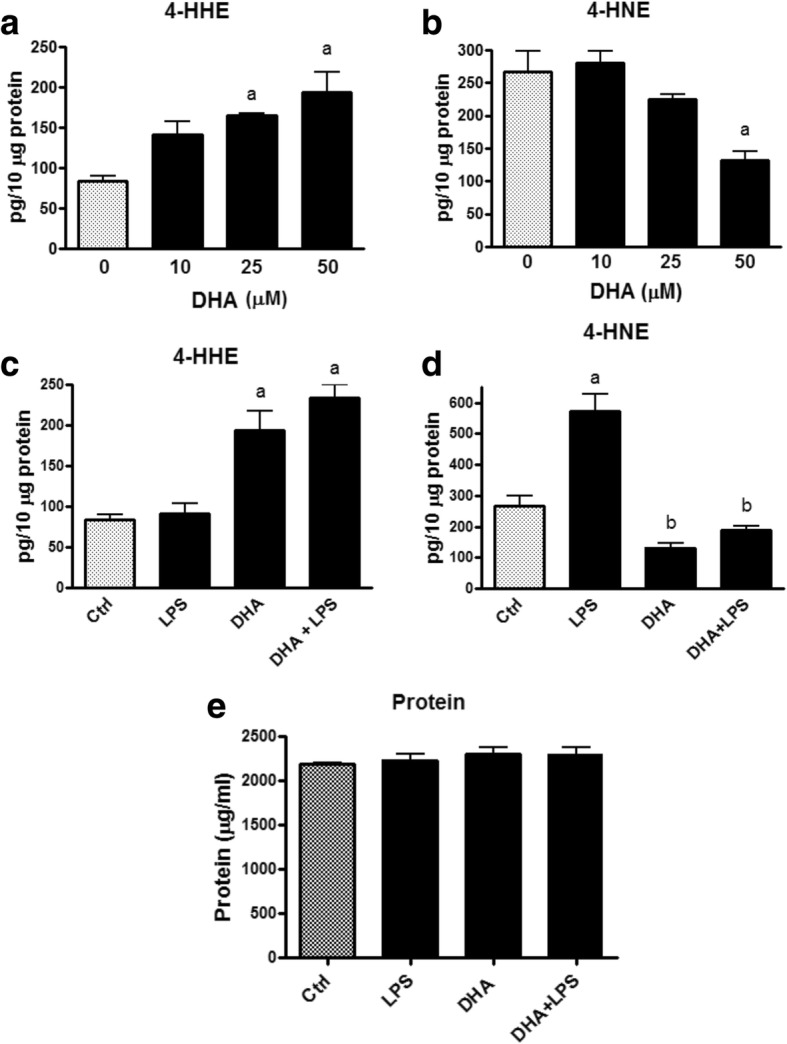
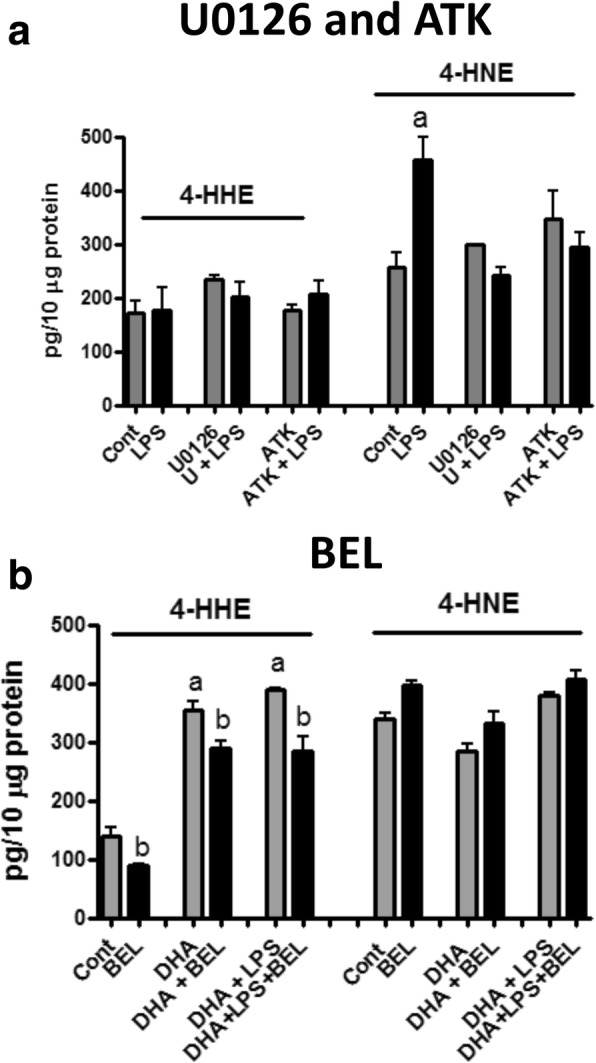
Results: DHA (12.5-100 μM), 4-HHE (1.25-10 μM), and 4-HNE (1.25-10 μM) dose dependently suppressed LPS-induced production of NO, ROS, and as p-cPLA2 in BV-2 microglial cells. With the same concentrations, these compounds could enhance Nrf2 and HO-1 expression in these cells. Based on the estimated IC50 values, 4-HHE and 4-HNE were five- to tenfold more potent than DHA in inhibiting LPS-induced NO, ROS, and p-cPLA2. LC-MS/MS analysis indicated ability for DHA (10-50 μM) to increase levels of 4-HHE and attenuate levels of 4-HNE in BV-2 microglial cells. Stimulation of cells with LPS caused an increase in 4-HNE which could be abrogated by cPLA2 inhibitor. In contrast, bromoenol lactone (BEL), a specific inhibitor for the Ca2+-independent phospholipase A2 (iPLA2), could only partially suppress levels of 4-HHE induced by DHA or DHA + LPS.
Conclusions: This study demonstrated the ability of DHA and its lipid peroxidation products, namely, 4-HHE and 4-HNE at 1.25-10 μM, to enhance Nrf2/HO-1 and mitigate LPS-induced NO, ROS, and p-cPLA2 in BV-2 microglial cells. In addition, LC-MS/MS analysis of the levels of 4-HHE and 4-HNE in microglial cells demonstrates that increases in production of 4-HHE from DHA and 4-HNE from LPS are mediated by different mechanisms.
Keywords: 4-Hydroxyhexenal (4-HHE); 4-Hydroxynonenal (4-HNE); Docosahexaenoic acid (DHA); HO-1; Lipopolysaccharide (LPS); Microglia; NO; Nrf2; ROS; cPLA2; iPLA2.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Paterniti I, Impellizzeri D, Di Paola R, Esposito E, Gladman S, Yip P, Priestley JV, Michael-Titus AT, Cuzzocrea S. Docosahexaenoic acid attenuates the early inflammatory response following spinal cord injury in mice: in-vivo and in-vitro studies. J Neuroinflammation. 2014;11:6. doi: 10.1186/1742-2094-11-6. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
