

Nitric oxide promotes cancer cell dedifferentiation by disrupting an Oct4:caveolin-1 complex: A new regulatory mechanism for cancer stem cell formation
- PMID: 29986880
- PMCID: PMC6120192
- DOI: 10.1074/jbc.RA117.000287
Nitric oxide promotes cancer cell dedifferentiation by disrupting an Oct4:caveolin-1 complex: A new regulatory mechanism for cancer stem cell formation
Abstract
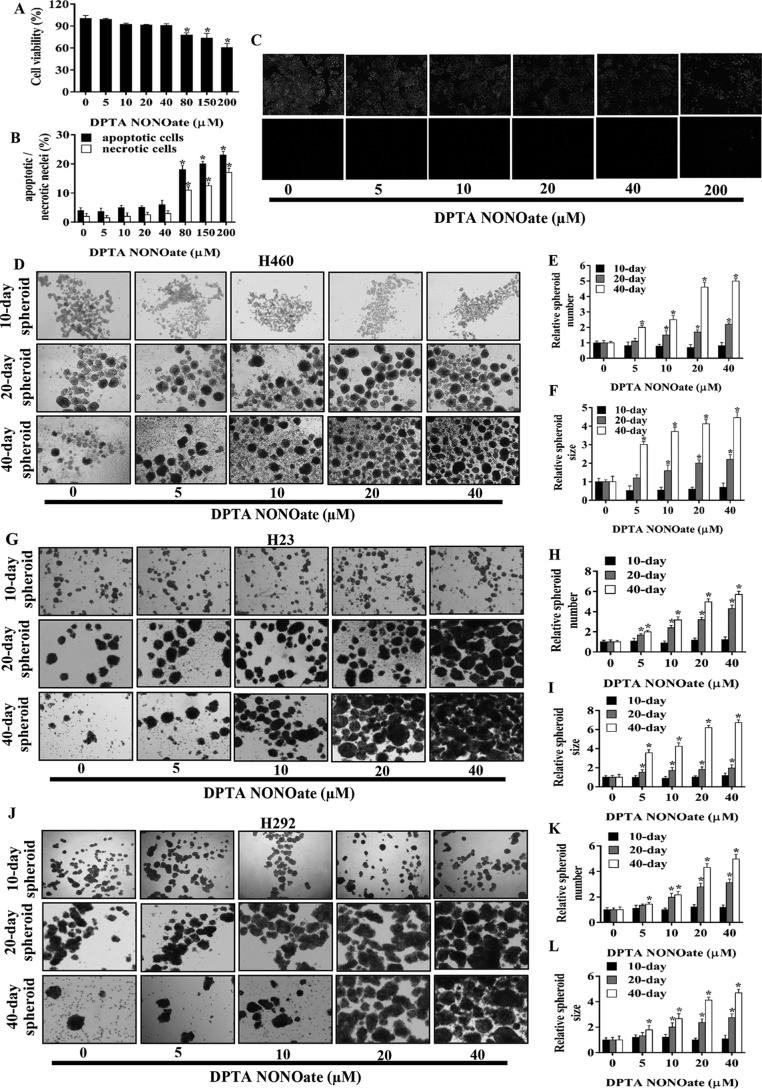
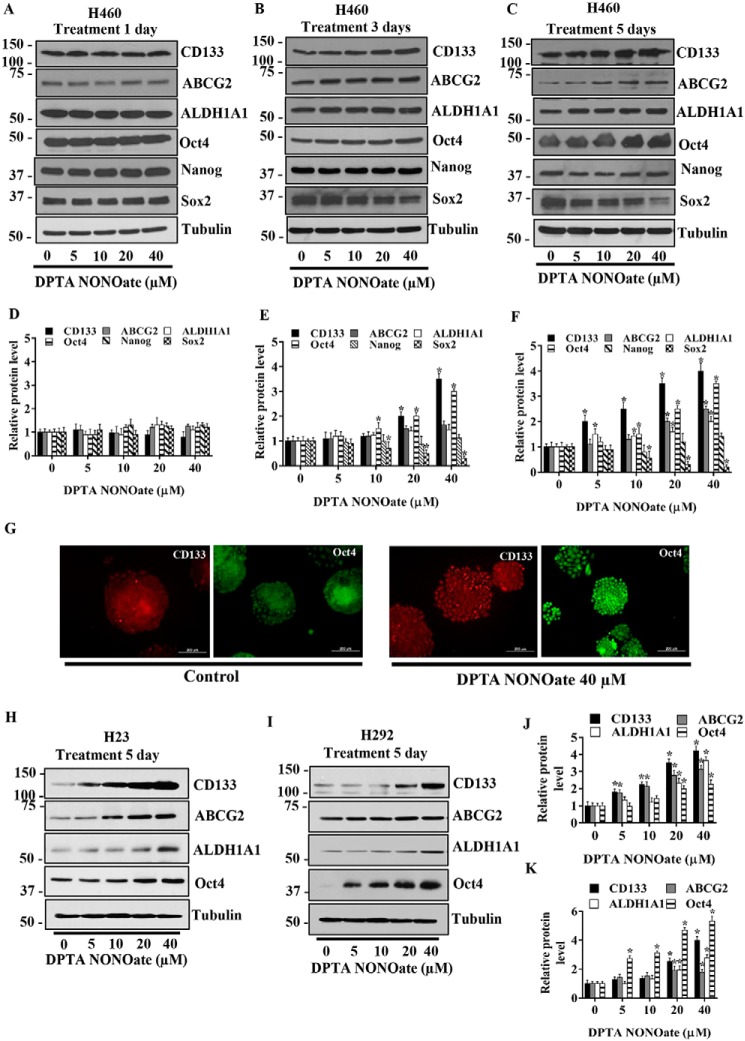
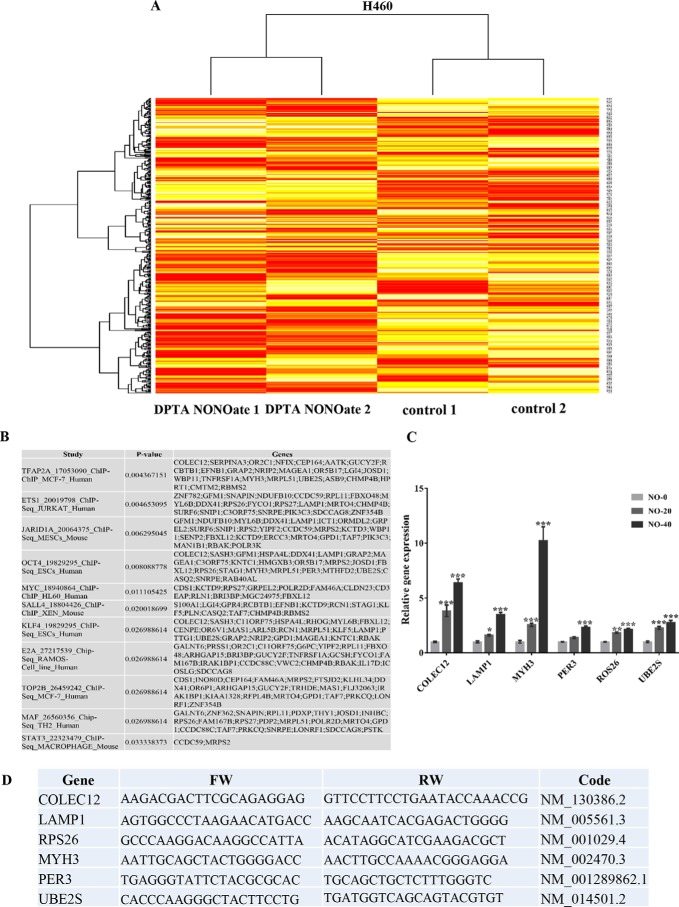
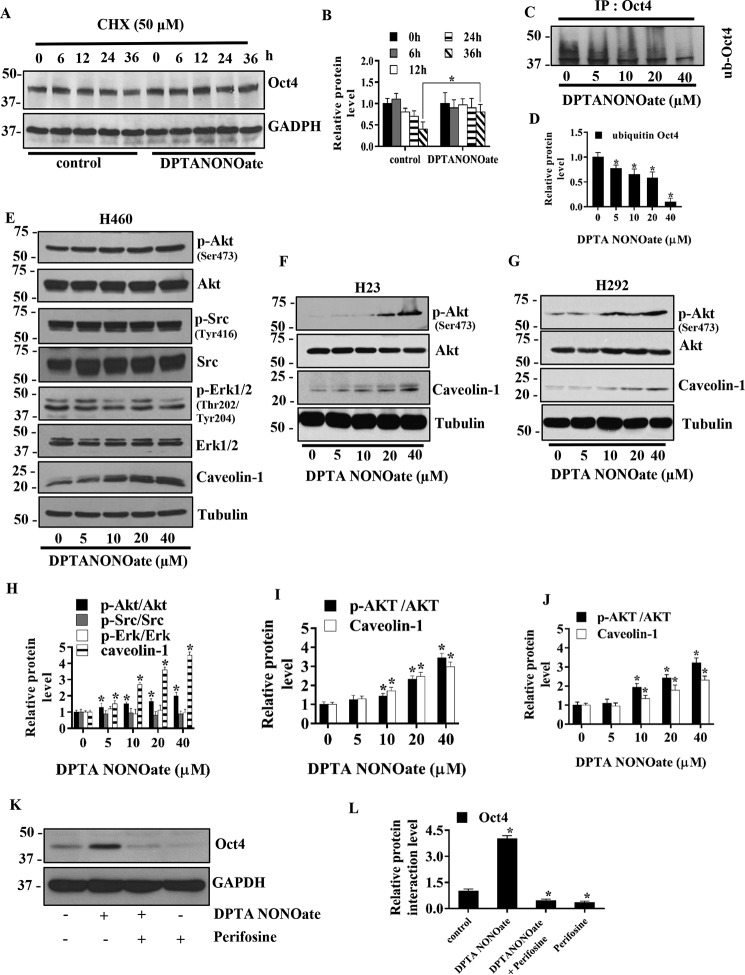
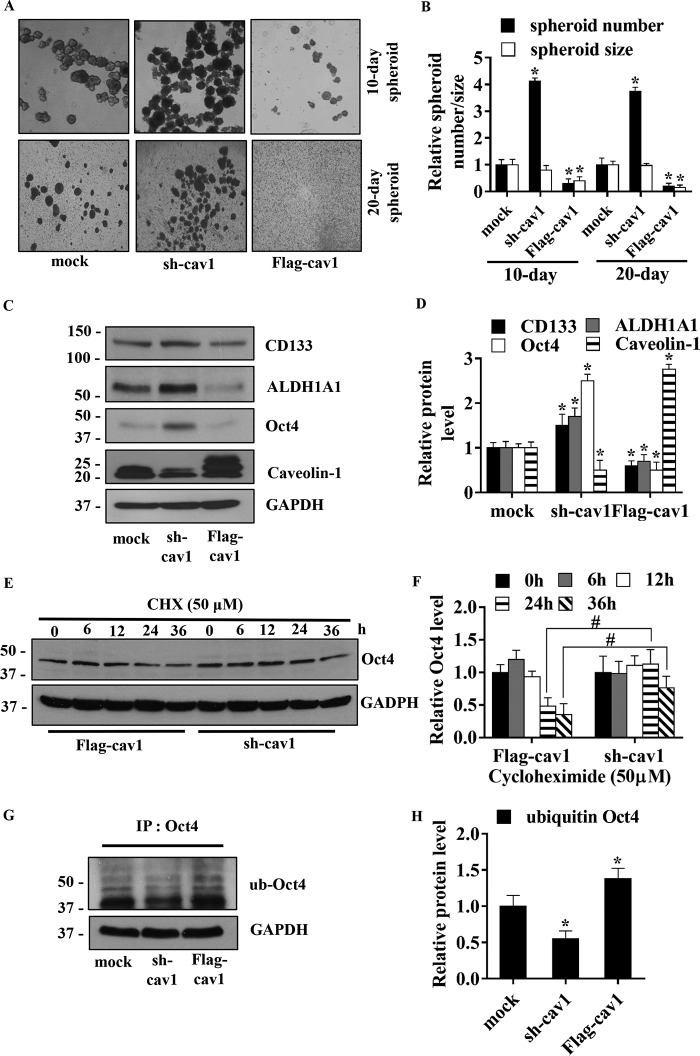
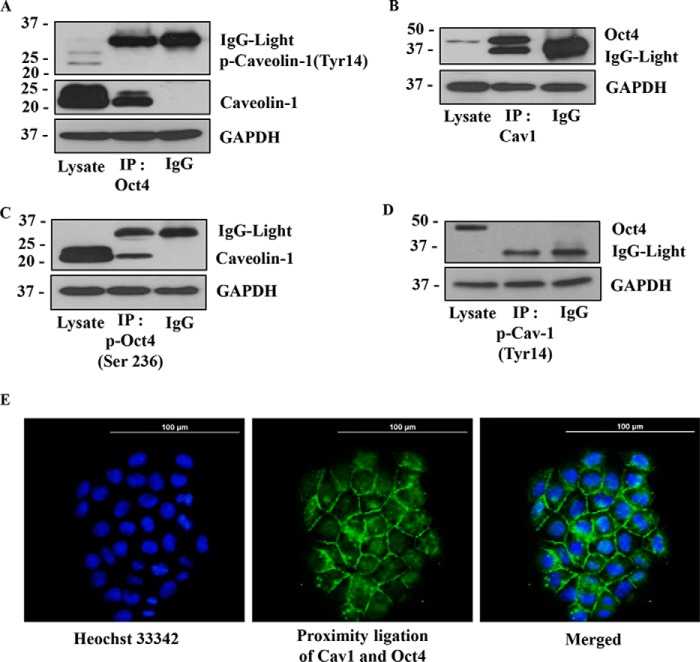
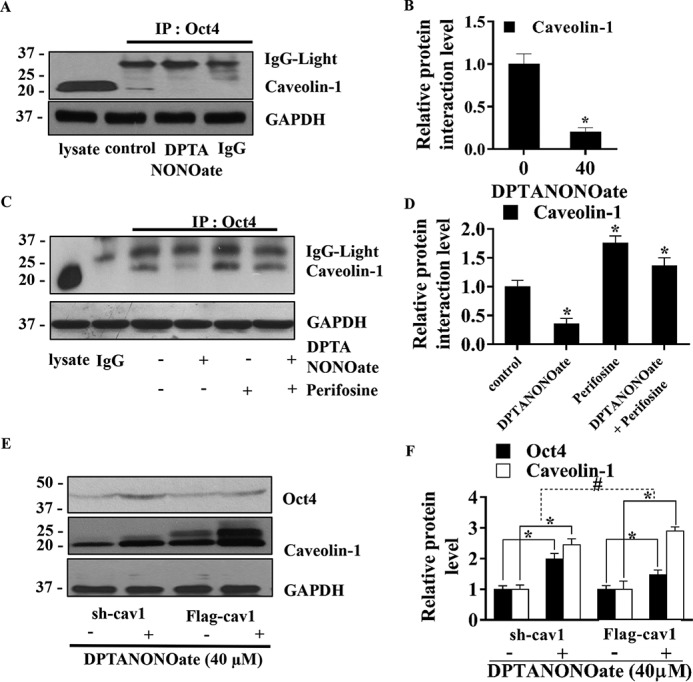
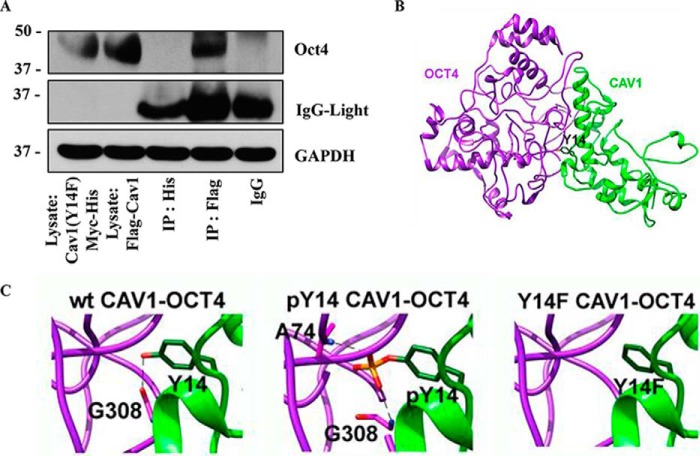
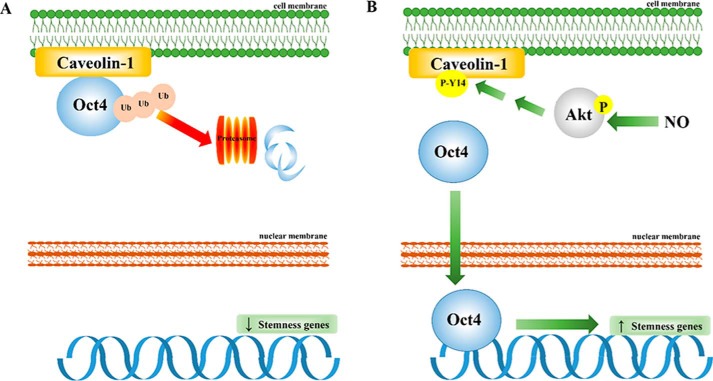
Cancer stem cells (CSCs) are unique populations of cells that can self-renew and generate different cancer cell lineages. Although CSCs are believed to be a promising target for novel therapies, the specific mechanisms by which these putative therapeutics could intervene are less clear. Nitric oxide (NO) is a biological mediator frequently up-regulated in tumors and has been linked to cancer aggressiveness. Here, we search for targets of NO that could explain its activity. We find that it directly affects the stability and function of octamer-binding transcription factor 4 (Oct4), known to drive the stemness of lung cancer cells. We demonstrated that NO promotes the CSC-regulatory activity of Oct4 through a mechanism that involves complex formation between Oct4 and the scaffolding protein caveolin-1 (Cav-1). In the absence of NO, Oct4 forms a molecular complex with Cav-1, which promotes the ubiquitin-mediated proteasomal degradation of Oct4. NO promotes Akt-dependent phosphorylation of Cav-1 at tyrosine 14, disrupting the Cav-1:Oct4 complex. Site-directed mutagenesis and computational modeling studies revealed that the hydroxyl moiety at tyrosine 14 of Cav-1 is crucial for its interaction with Oct4. Both removal of the hydroxyl via mutation to phenylalanine and phosphorylation lead to an increase in binding free energy (ΔGbind) between Oct4 and Cav-1, destabilizing the complex. Together, these results unveiled a novel mechanism of CSC regulation through NO-mediated stabilization of Oct4, a key stem cell transcription factor, and point to new opportunities to design CSC-related therapeutics.
Keywords: GSK3β; OCT4; cancer biology; cancer stem cells; caveolin; cell signaling; differentiation; lung cancer; nitric oxide, caveolin-1; proteasome; protein degradation; protein–protein interaction; regulation.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
