

Conserved motifs on the cytoplasmic face of the protein translocation channel are critical for the transition between resting and active conformations
- PMID: 29986881
- PMCID: PMC6120202
- DOI: 10.1074/jbc.RA118.004123
Conserved motifs on the cytoplasmic face of the protein translocation channel are critical for the transition between resting and active conformations
Abstract
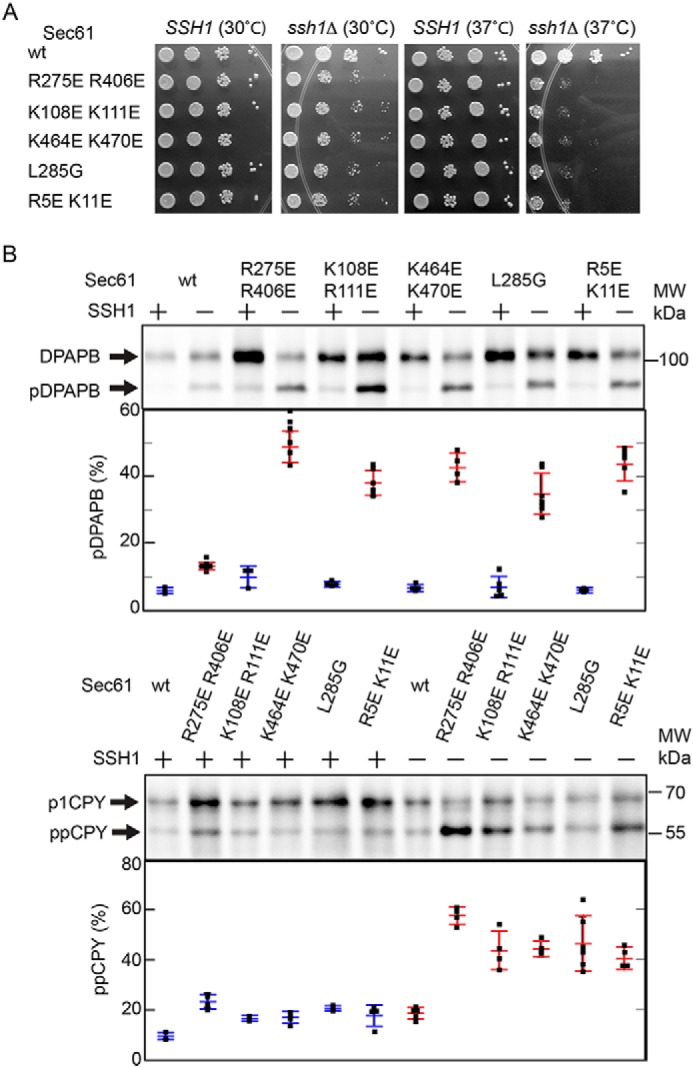
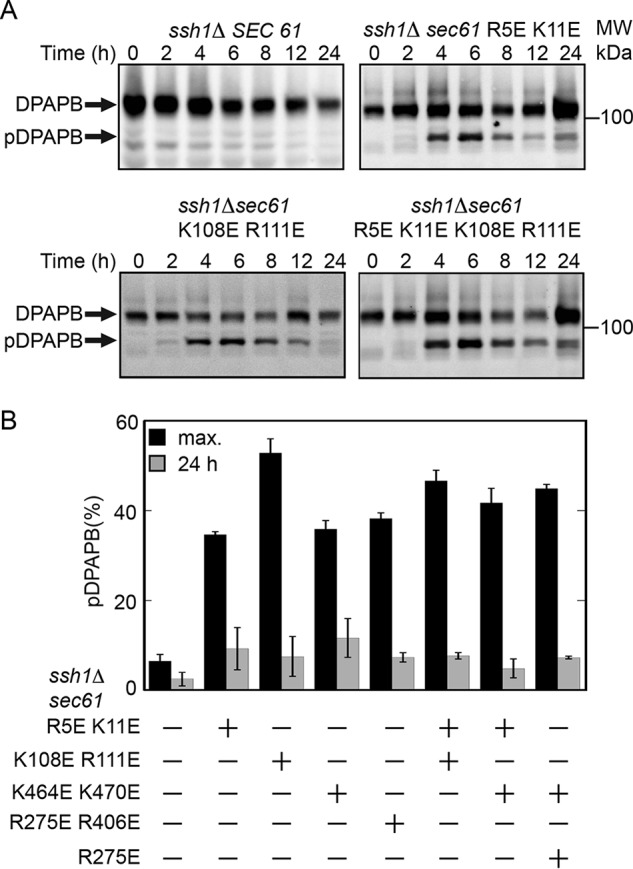
The Sec61 complex is the primary cotranslational protein translocation channel in yeast (Saccharomyces cerevisiae). The structural transition between the closed inactive conformation of the Sec61 complex and its open and active conformation is thought to be promoted by binding of the ribosome nascent-chain complex to the cytoplasmic surface of the Sec61 complex. Here, we have analyzed new yeast Sec61 mutants that selectively interfere with cotranslational translocation across the endoplasmic reticulum. We found that a single substitution at the junction between transmembrane segment TM7 and the L6/7 loop interferes with cotranslational translocation by uncoupling ribosome binding to the L6/7 loop from the separation of the lateral gate transmembrane spans. Substitutions replacing basic residues with acidic residues in the C-terminal tail of Sec61 had an unanticipated impact upon binding of ribosomes to the Sec61 complex. We found that similar charge-reversal mutations in the N-terminal tail and in cytoplasmic loop L2/3 did not alter ribosome binding but interfered with translocation channel gating. These findings indicated that these segments are important for the structural transition between the inactive and active conformations of the Sec61 complex. In summary our results have identified additional cytosolic segments of the Sec61 complex important for promoting the structural transition between the closed and open conformations of the complex. We conclude that positively charged residues in multiple cytosolic segments, as well as bulky hydrophobic residues in the L6/7-TM7 junction, are required for cotranslational translocation or integration of membrane proteins by the Sec61 complex.
Keywords: Sec61; endoplasmic reticulum (ER); membrane protein; protein synthesis; protein translocation; ribosome; translocation channel.
© 2018 Mandon et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
