

Protein moonlighting elucidates the essential human pathway catalyzing lipoic acid assembly on its cognate enzymes
- PMID: 29987032
- PMCID: PMC6064980
- DOI: 10.1073/pnas.1805862115
Protein moonlighting elucidates the essential human pathway catalyzing lipoic acid assembly on its cognate enzymes
Abstract
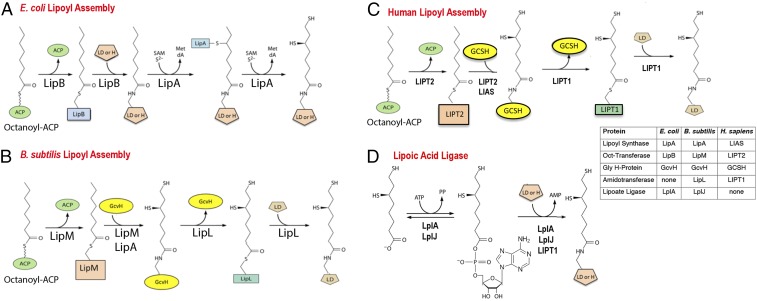
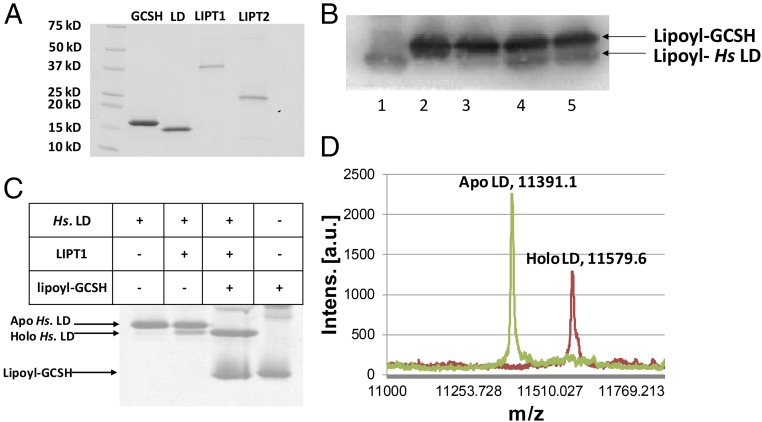
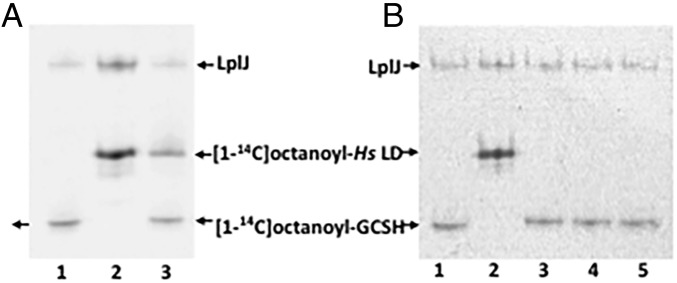
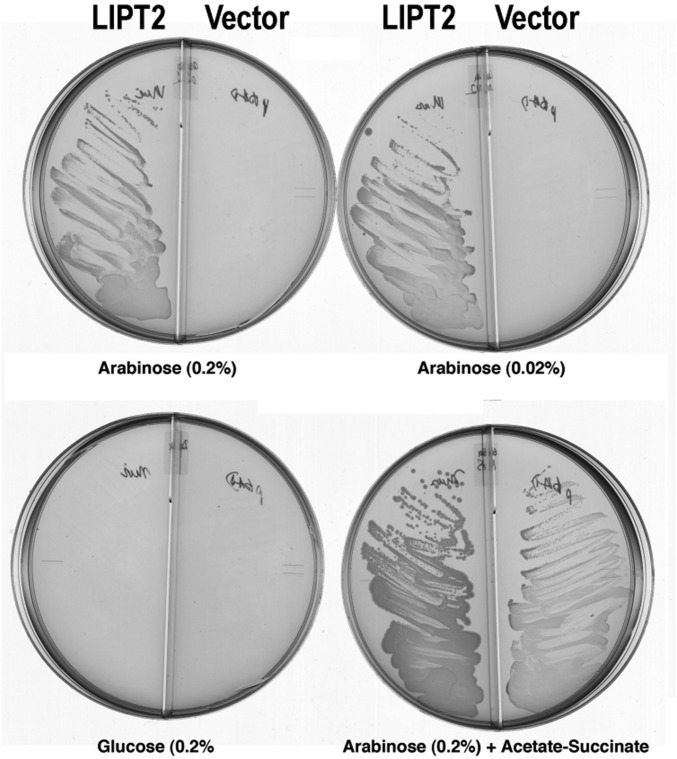
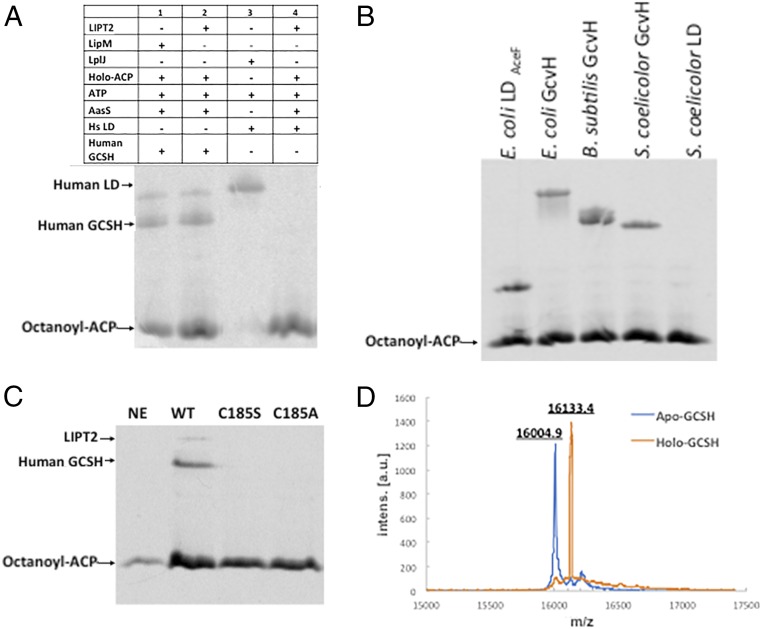
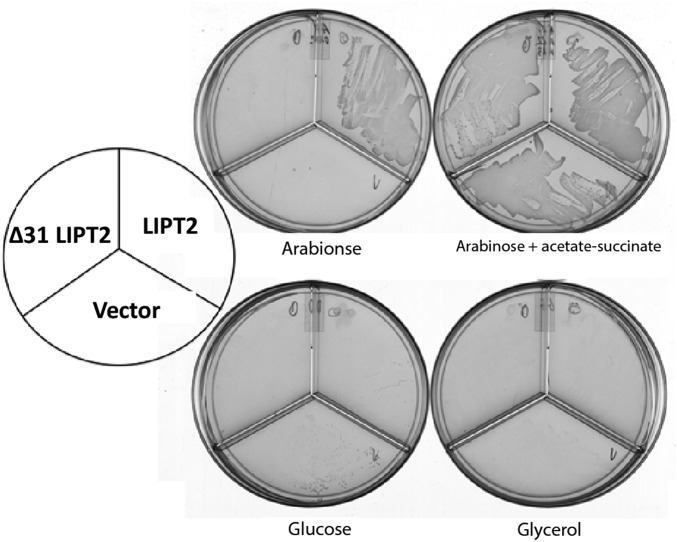
The lack of attachment of lipoic acid to its cognate enzyme proteins results in devastating human metabolic disorders. These mitochondrial disorders are evident soon after birth and generally result in early death. The mutations causing specific defects in lipoyl assembly map in three genes, LIAS, LIPT1, and LIPT2 Although physiological roles have been proposed for the encoded proteins, only the LIPT1 protein had been studied at the enzyme level. LIPT1 was reported to catalyze only the second partial reaction of the classical lipoate ligase mechanism. We report that the physiologically relevant LIPT1 enzyme activity is transfer of lipoyl moieties from the H protein of the glycine cleavage system to the E2 subunits of the 2-oxoacid dehydrogenases required for respiration (e.g., pyruvate dehydrogenase) and amino acid degradation. We also report that LIPT2 encodes an octanoyl transferase that initiates lipoyl group assembly. The human pathway is now biochemically defined.
Keywords: 2-oxoacid dehydrogenases; glycine cleavage system; inborn errors; lipoic acid; mitochondrial disorder.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Reed LJ. A trail of research from lipoic acid to alpha-keto acid dehydrogenase complexes. J Biol Chem. 2001;276:38329–38336. - PubMed
-
- Reed LJ, DeBUSK BG, Gunsalus IC, Hornberger CS., Jr Crystalline alpha-lipoic acid; a catalytic agent associated with pyruvate dehydrogenase. Science. 1951;114:93–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
