

Scleral hypoxia is a target for myopia control
- PMID: 29987045
- PMCID: PMC6064999
- DOI: 10.1073/pnas.1721443115
Scleral hypoxia is a target for myopia control
Abstract
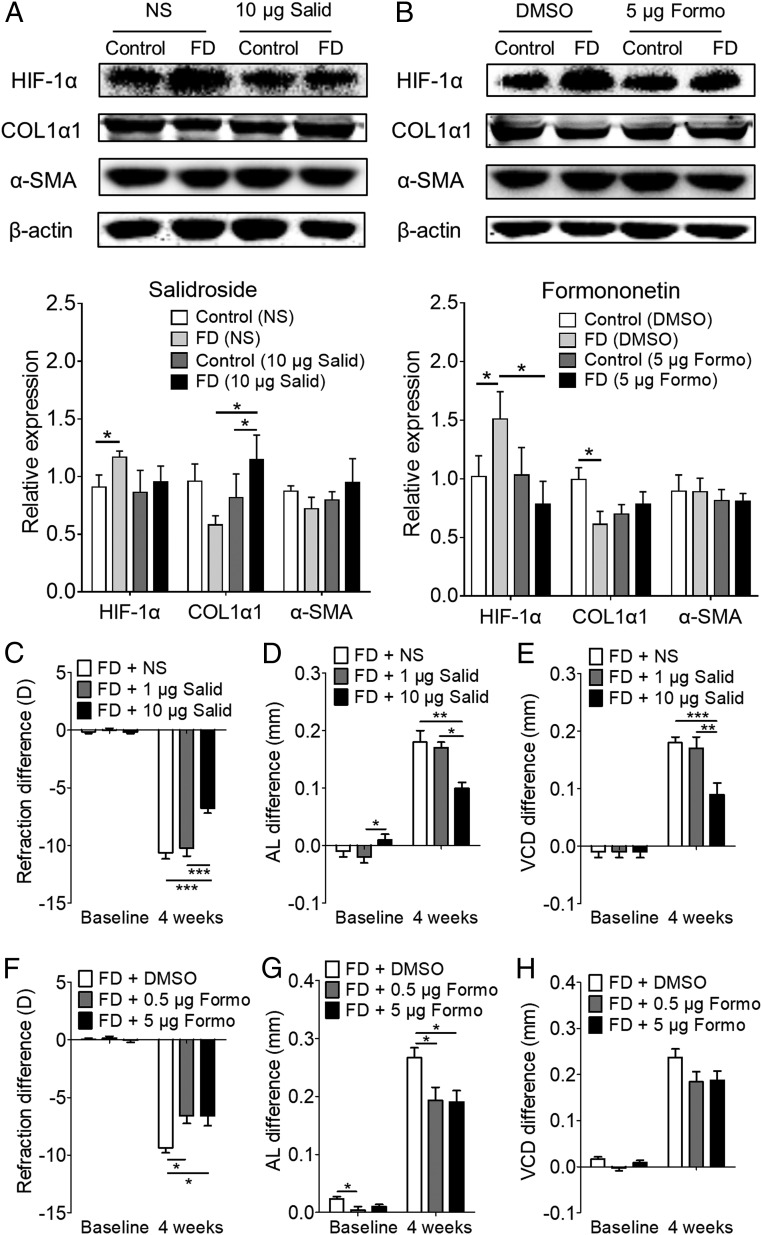
Worldwide, myopia is the leading cause of visual impairment. It results from inappropriate extension of the ocular axis and concomitant declines in scleral strength and thickness caused by extracellular matrix (ECM) remodeling. However, the identities of the initiators and signaling pathways that induce scleral ECM remodeling in myopia are unknown. Here, we used single-cell RNA-sequencing to identify pathways activated in the sclera during myopia development. We found that the hypoxia-signaling, the eIF2-signaling, and mTOR-signaling pathways were activated in murine myopic sclera. Consistent with the role of hypoxic pathways in mouse model of myopia, nearly one third of human myopia risk genes from the genome-wide association study and linkage analyses interact with genes in the hypoxia-inducible factor-1α (HIF-1α)-signaling pathway. Furthermore, experimental myopia selectively induced HIF-1α up-regulation in the myopic sclera of both mice and guinea pigs. Additionally, hypoxia exposure (5% O2) promoted myofibroblast transdifferentiation with down-regulation of type I collagen in human scleral fibroblasts. Importantly, the antihypoxia drugs salidroside and formononetin down-regulated HIF-1α expression as well as the phosphorylation levels of eIF2α and mTOR, slowing experimental myopia progression without affecting normal ocular growth in guinea pigs. Furthermore, eIF2α phosphorylation inhibition suppressed experimental myopia, whereas mTOR phosphorylation induced myopia in normal mice. Collectively, these findings defined an essential role of hypoxia in scleral ECM remodeling and myopia development, suggesting a therapeutic approach to control myopia by ameliorating hypoxia.
Keywords: HIF-1α; myopia; scRNA-seq; scleral ECM remodeling; scleral hypoxia.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






References
-
- Dolgin E. The myopia boom. Nature. 2015;519:276–278. - PubMed
-
- Holden BA, et al. Global prevalence of myopia and high myopia and temporal trends from 2000 through 2050. Ophthalmology. 2016;123:1036–1042. - PubMed
-
- Flitcroft DI. The complex interactions of retinal, optical and environmental factors in myopia aetiology. Prog Retin Eye Res. 2012;31:622–660. - PubMed
-
- Leo SW. Scientific Bureau of World Society of Paediatric Ophthalmology and Strabismus (WSPOS) Current approaches to myopia control. Curr Opin Ophthalmol. 2017;28:267–275. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
