

Oncogenic addiction to high 26S proteasome level
- PMID: 29991718
- PMCID: PMC6039477
- DOI: 10.1038/s41419-018-0806-4
Oncogenic addiction to high 26S proteasome level
Abstract
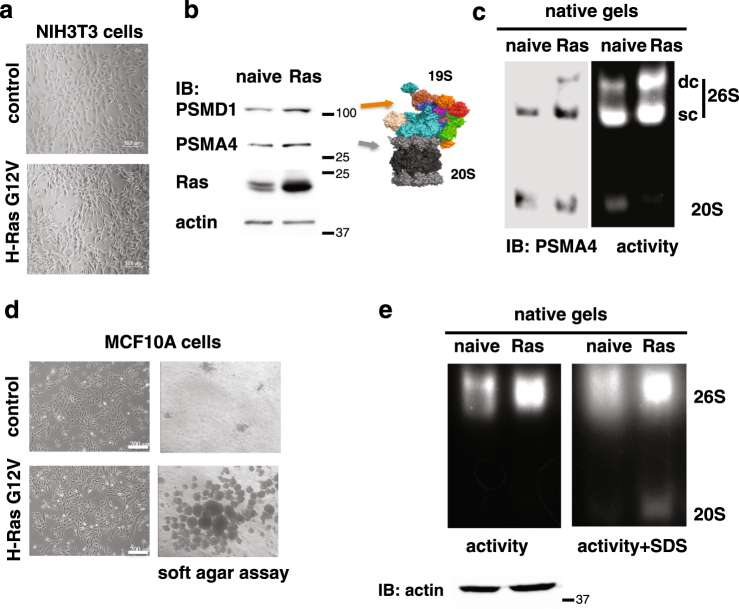
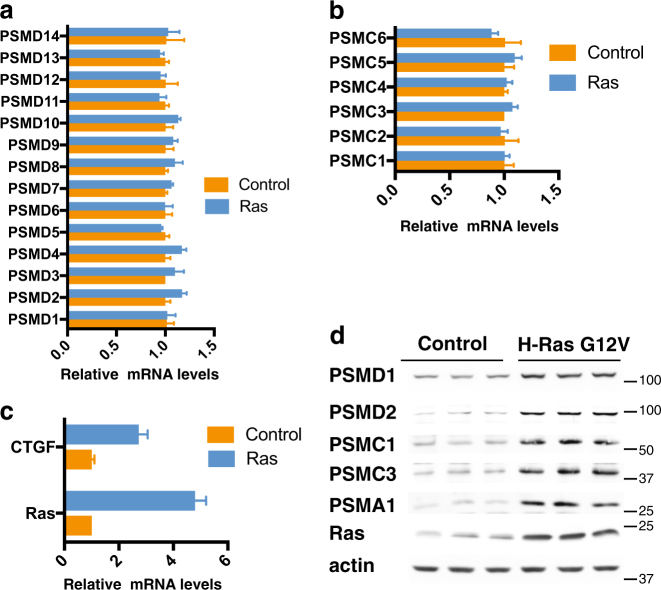
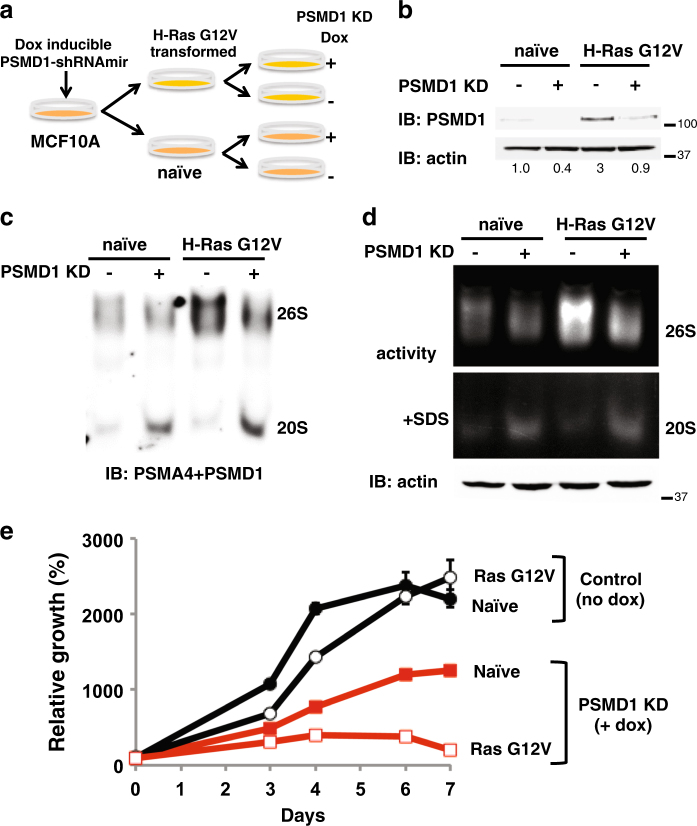
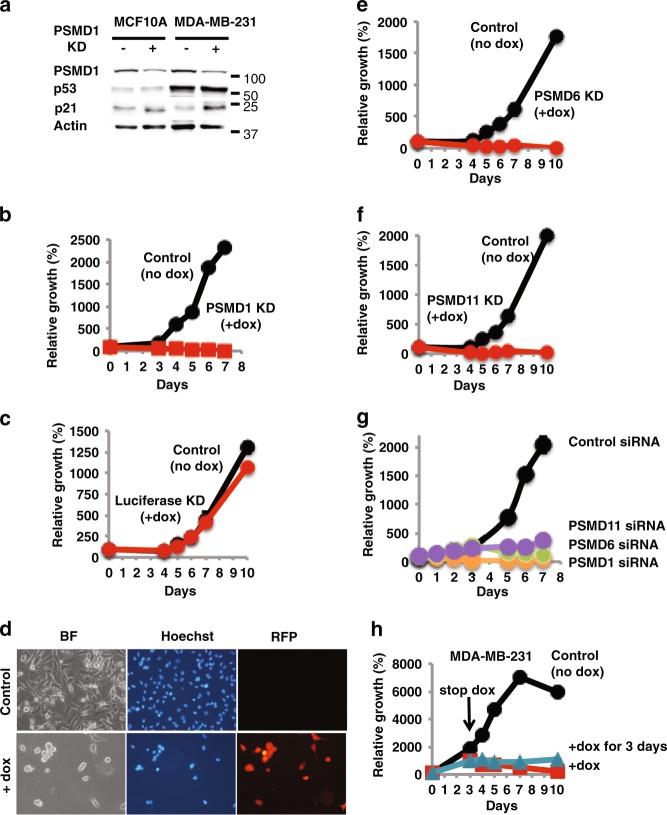
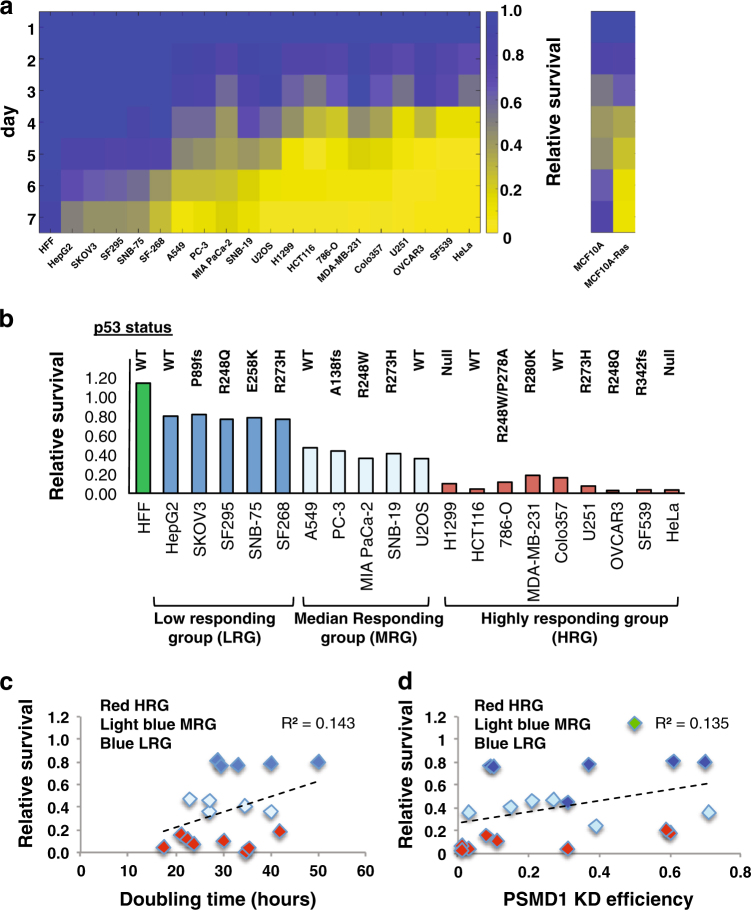
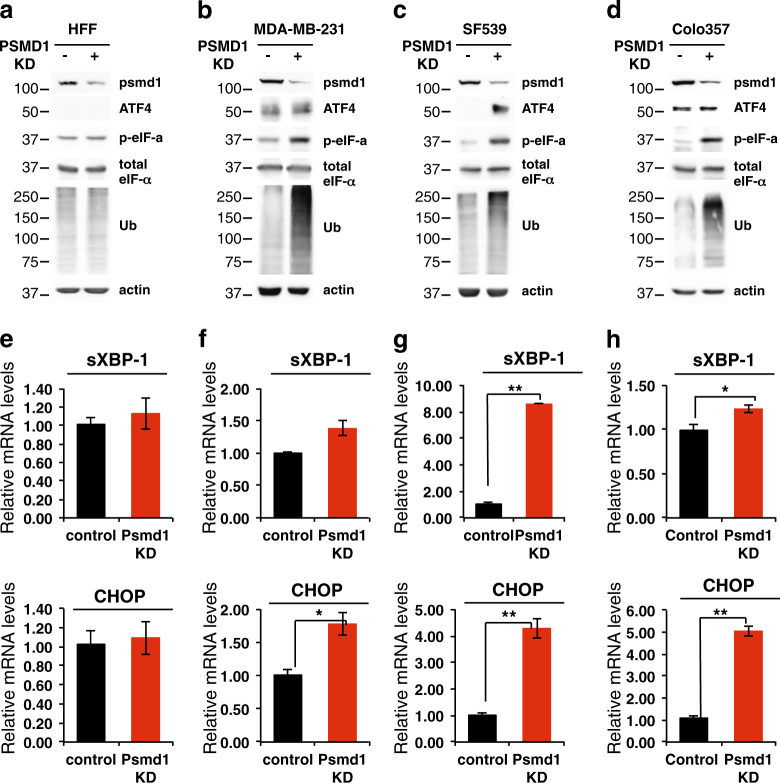
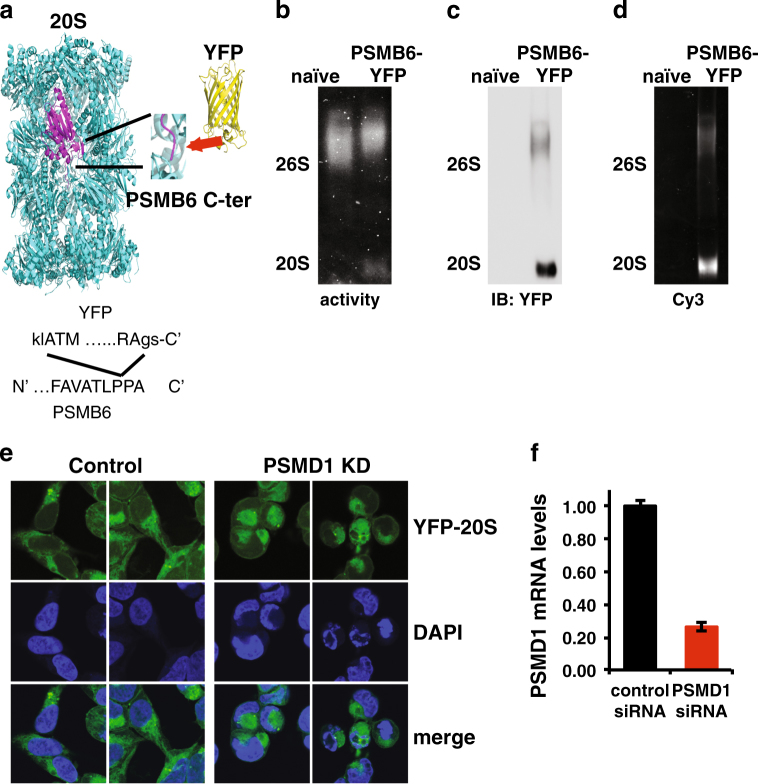
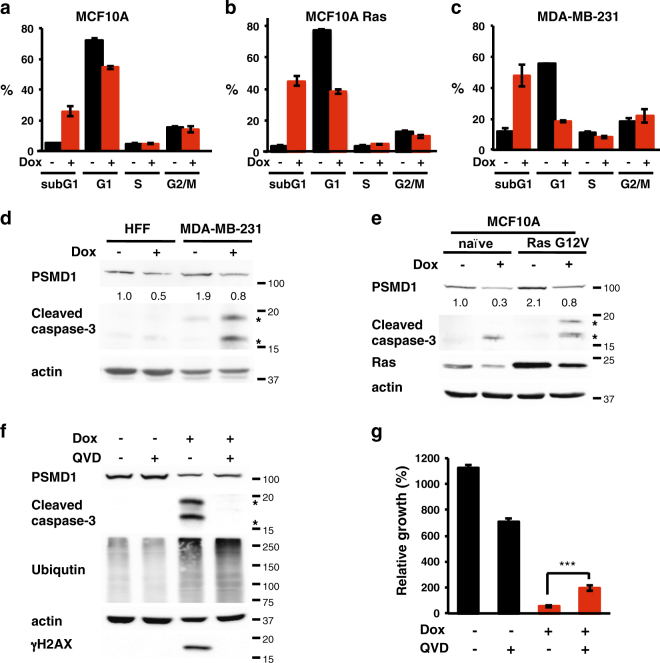
Proteasomes are large intracellular complexes responsible for the degradation of cellular proteins. The altered protein homeostasis of cancer cells results in increased dependency on proteasome function. The cellular proteasome composition comprises the 20S catalytic complex that is frequently capped with the 19S regulatory particle in forming the 26S proteasome. Proteasome inhibitors target the catalytic barrel (20S) and thus this inhibition does not allow the deconvolution of the distinct roles of 20S versus 26S proteasomes in cancer progression. We examined the degree of dependency of cancer cells specifically to the level of the 26S proteasome complex. Oncogenic transformation of human and mouse immortalized cells with mutant Ras induced a strong posttranscriptional increase of the 26S proteasome subunits, giving rise to high 26S complex levels. Depletion of a single subunit of the 19S RP was sufficient to reduce the 26S proteasome level and lower the cellular 26S/20S ratio. Under this condition the viability of the Ras-transformed MCF10A cells was severely compromised. This observation led us to hypothesize that cancer cell survival is dependent on maximal utilization of its 26S proteasomes. We validated this possibility in a large number of cancer cell lines and found that partial reduction of the 26S proteasome level impairs viability in all cancer cells examined and was not correlated with cell doubling time or reduction efficiency. Interstingly, normal human fibroblasts are refractory to the same type of 26S proteasome reduction. The suppression of 26S proteasomes in cancer cells activated the UPR and caspase-3 and cells stained positive with Annexin V. In addition, suppression of the 26S proteasome resulted in cellular proteasome redistribution, cytoplasm shrinkage, and nuclear deformation, the hallmarks of apoptosis. The observed tumor cell-specific addiction to the 26S proteasome levels sets the stage for future strategies in exploiting this dependency in cancer therapy.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Depleting the 19S proteasome regulatory PSMD1 subunit as a cancer therapy strategy.Cancer Med. 2023 May;12(9):10781-10790. doi: 10.1002/cam4.5775. Epub 2023 Mar 19. Cancer Med. 2023. PMID: 36934426 Free PMC article.
-
Method of Monitoring 26S Proteasome in Cells Revealed the Crucial Role of PSMA3 C-Terminus in 26S Integrity.Biomolecules. 2023 Jun 15;13(6):992. doi: 10.3390/biom13060992. Biomolecules. 2023. PMID: 37371572 Free PMC article.
-
Mapping the murine cardiac 26S proteasome complexes.Circ Res. 2006 Aug 18;99(4):362-71. doi: 10.1161/01.RES.0000237386.98506.f7. Epub 2006 Jul 20. Circ Res. 2006. PMID: 16857966
-
Proteasomes of the yeast S. cerevisiae: genes, structure and functions.Mol Biol Rep. 1995;21(1):3-10. doi: 10.1007/BF00990964. Mol Biol Rep. 1995. PMID: 7565661 Review.
-
Regulation of proteasome complexes by gamma-interferon and phosphorylation.Biochimie. 2001 Mar-Apr;83(3-4):363-6. doi: 10.1016/s0300-9084(01)01249-4. Biochimie. 2001. PMID: 11295498 Review.
Cited by
-
Endocrine Resistance in Breast Cancer: The Role of Estrogen Receptor Stability.Cells. 2020 Sep 11;9(9):2077. doi: 10.3390/cells9092077. Cells. 2020. PMID: 32932819 Free PMC article. Review.
-
PSMD1 as a prognostic marker and potential target in oropharyngeal cancer.BMC Cancer. 2023 Dec 16;23(1):1242. doi: 10.1186/s12885-023-11689-2. BMC Cancer. 2023. PMID: 38104103 Free PMC article.
-
POH1/Rpn11/PSMD14: a journey from basic research in fission yeast to a prognostic marker and a druggable target in cancer cells.Br J Cancer. 2022 Sep;127(5):788-799. doi: 10.1038/s41416-022-01829-z. Epub 2022 May 2. Br J Cancer. 2022. PMID: 35501388 Free PMC article. Review.
-
Evaluation of the NRF1-proteasome axis as a therapeutic target in breast cancer.Sci Rep. 2023 Sep 22;13(1):15843. doi: 10.1038/s41598-023-43121-x. Sci Rep. 2023. PMID: 37739987 Free PMC article.
-
PSMD7 downregulation suppresses lung cancer progression by regulating the p53 pathway.J Cancer. 2021 Jun 11;12(16):4945-4957. doi: 10.7150/jca.53613. eCollection 2021. J Cancer. 2021. PMID: 34234864 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
