

NaV1.4 DI-S4 periodic paralysis mutation R222W enhances inactivation and promotes leak current to attenuate action potentials and depolarize muscle fibers
- PMID: 29991727
- PMCID: PMC6039468
- DOI: 10.1038/s41598-018-28594-5
NaV1.4 DI-S4 periodic paralysis mutation R222W enhances inactivation and promotes leak current to attenuate action potentials and depolarize muscle fibers
Abstract
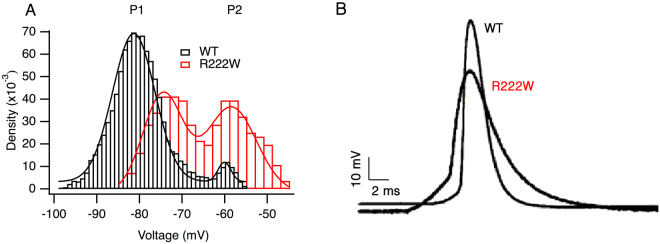
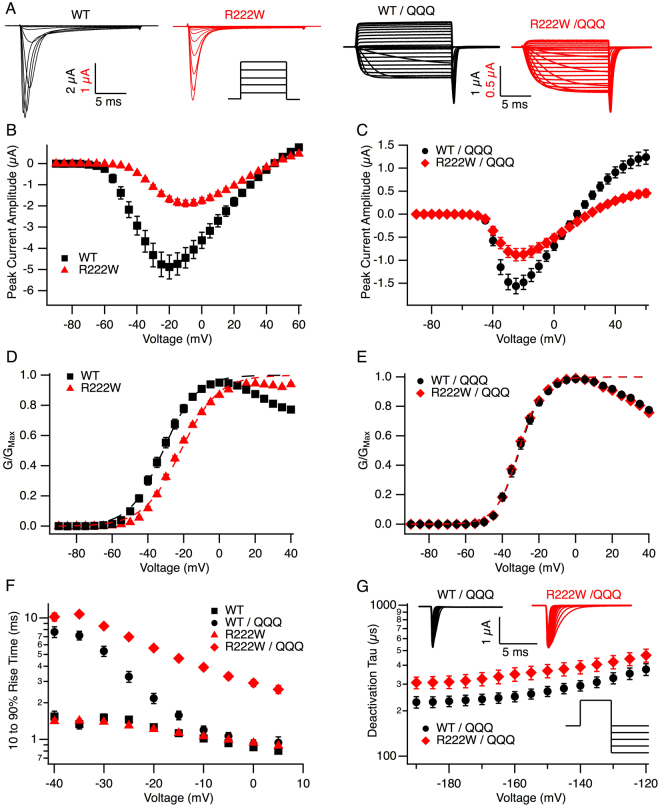
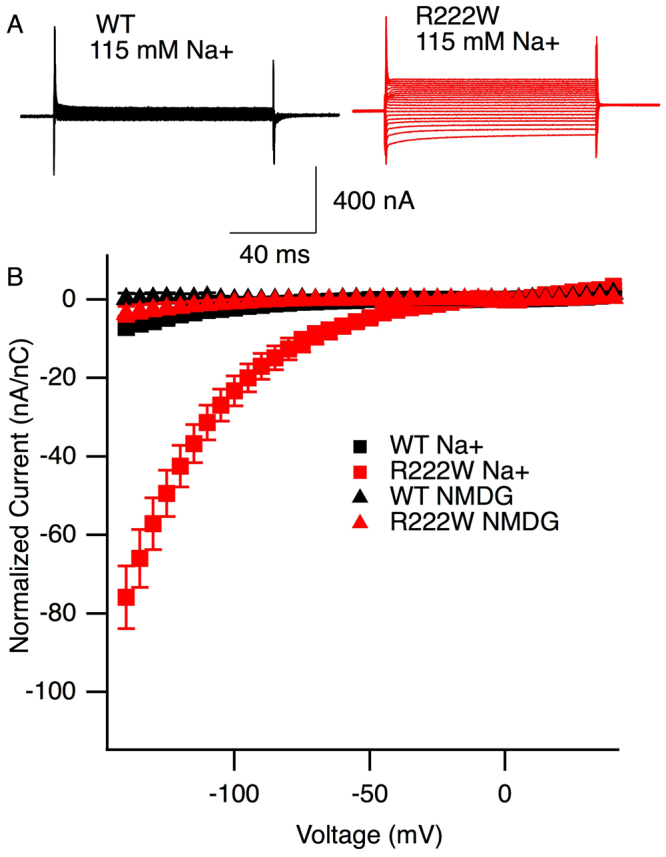
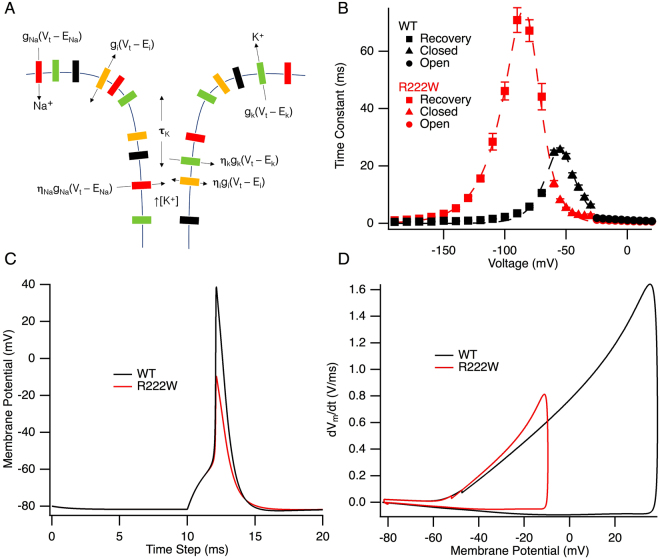
Hypokalemic periodic paralysis is a skeletal muscle disease characterized by episodic weakness associated with low serum potassium. We compared clinical and biophysical effects of R222W, the first hNaV1.4 domain I mutation linked to this disease. R222W patients exhibited a higher density of fibers with depolarized resting membrane potentials and produced action potentials that were attenuated compared to controls. Functional characterization of the R222W mutation in heterologous expression included the inactivation deficient IFM/QQQ background to isolate activation. R222W decreased sodium current and slowed activation without affecting probability. Consistent with the phenotype of muscle weakness, R222W shifted fast inactivation to hyperpolarized potentials, promoted more rapid entry, and slowed recovery. R222W increased the extent of slow inactivation and slowed its recovery. A two-compartment skeletal muscle fiber model revealed that defects in fast inactivation sufficiently explain action potential attenuation in patients. Molecular dynamics simulations showed that R222W disrupted electrostatic interactions within the gating pore, supporting the observation that R222W promotes omega current at hyperpolarized potentials. Sodium channel inactivation defects produced by R222W are the primary driver of skeletal muscle fiber action potential attenuation, while hyperpolarization-induced omega current produced by that mutation promotes muscle fiber depolarization.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Groome, J. R., Moreau, A. & Delemotte, L. Gating pore currents in sodium channels. In: Handbook of Experimental Pharmacology: Sodium channels and related disorders (ed. Chahine, M.) (Springer, Berlin, Heidelberg) (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
