

Image-Based Profiling of Synaptic Connectivity in Primary Neuronal Cell Culture
- PMID: 29997468
- PMCID: PMC6028601
- DOI: 10.3389/fnins.2018.00389
Image-Based Profiling of Synaptic Connectivity in Primary Neuronal Cell Culture
Abstract
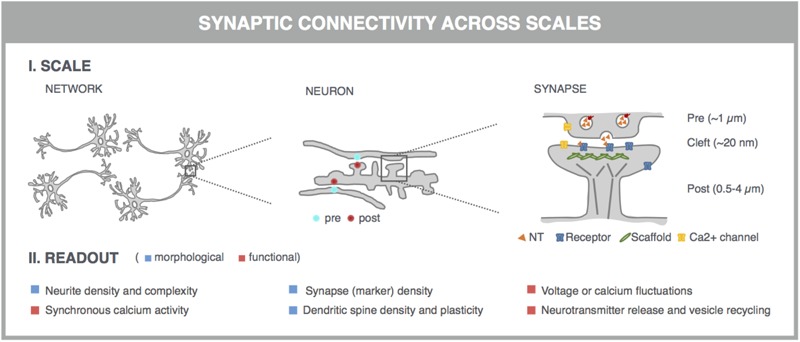
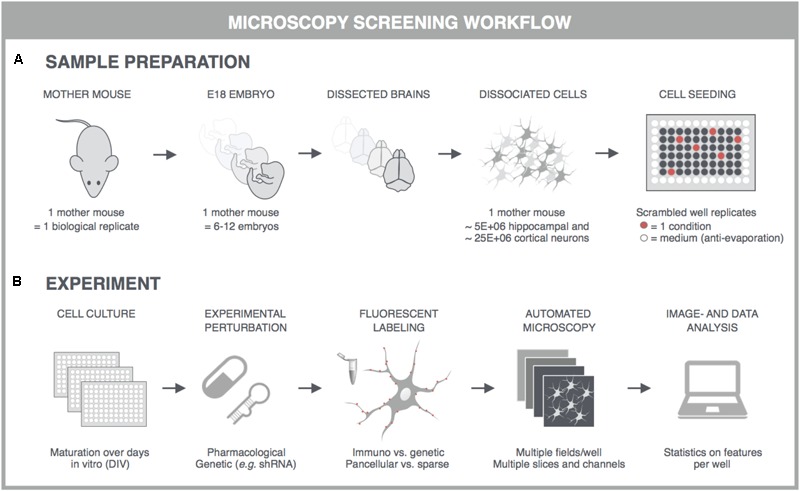
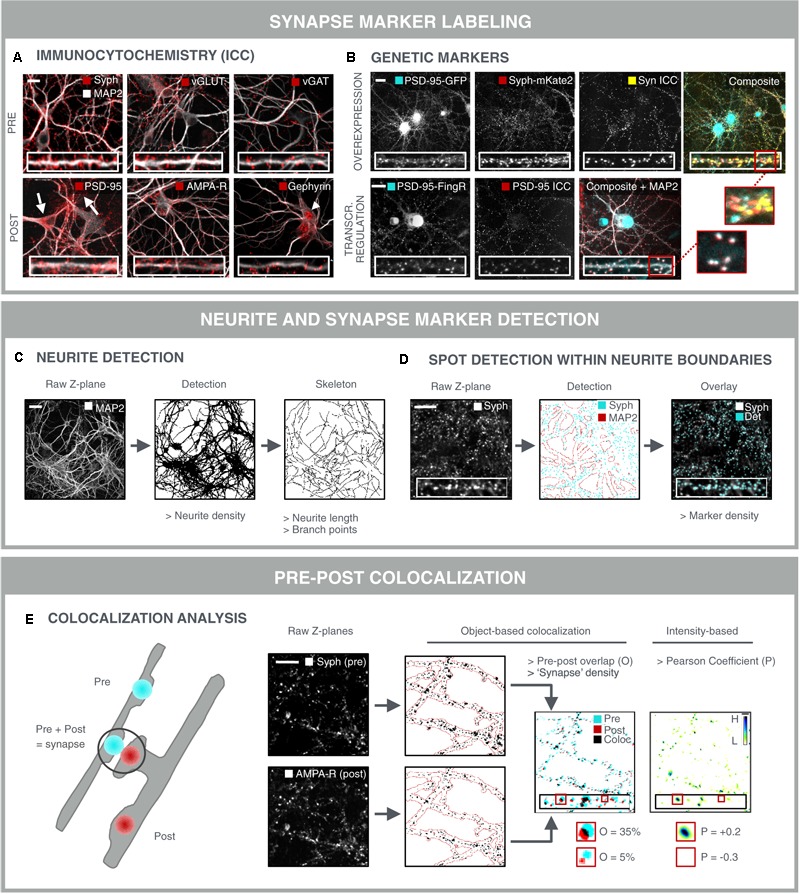
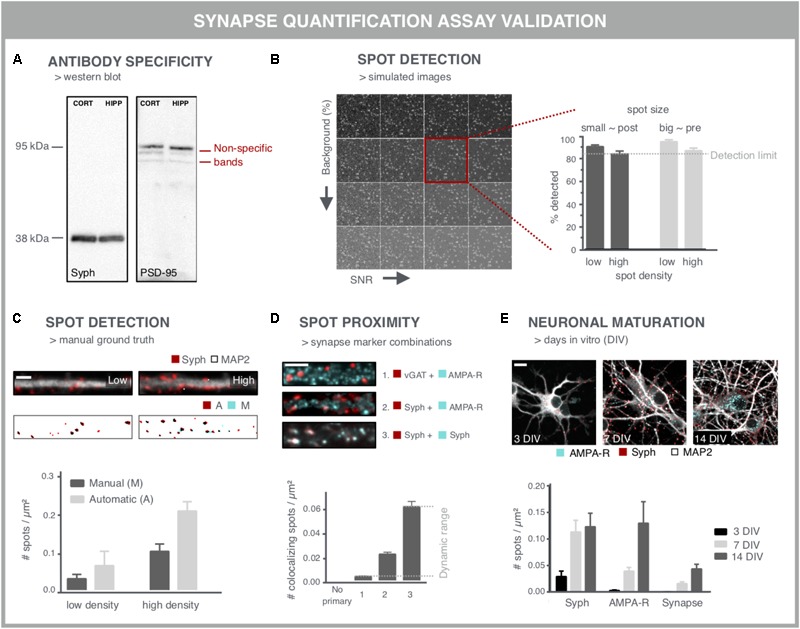
Neurological disorders display a broad spectrum of clinical manifestations. Yet, at the cellular level, virtually all these diseases converge into a common phenotype of dysregulated synaptic connectivity. In dementia, synapse dysfunction precedes neurodegeneration and cognitive impairment by several years, making the synapse a crucial entry point for the development of diagnostic and therapeutic strategies. Whereas high-resolution imaging and biochemical fractionations yield detailed insight into the molecular composition of the synapse, standardized assays are required to quickly gauge synaptic connectivity across large populations of cells under a variety of experimental conditions. Such screening capabilities have now become widely accessible with the advent of high-throughput, high-content microscopy. In this review, we discuss how microscopy-based approaches can be used to extract quantitative information about synaptic connectivity in primary neurons with deep coverage. We elaborate on microscopic readouts that may serve as a proxy for morphofunctional connectivity and we critically analyze their merits and limitations. Finally, we allude to the potential of alternative culture paradigms and integrative approaches to enable comprehensive profiling of synaptic connectivity.
Keywords: dendritic spine; fluorescent labeling; high-content screening; image analysis; morphofunctional connectivity; primary neuronal culture; synapse.
Figures
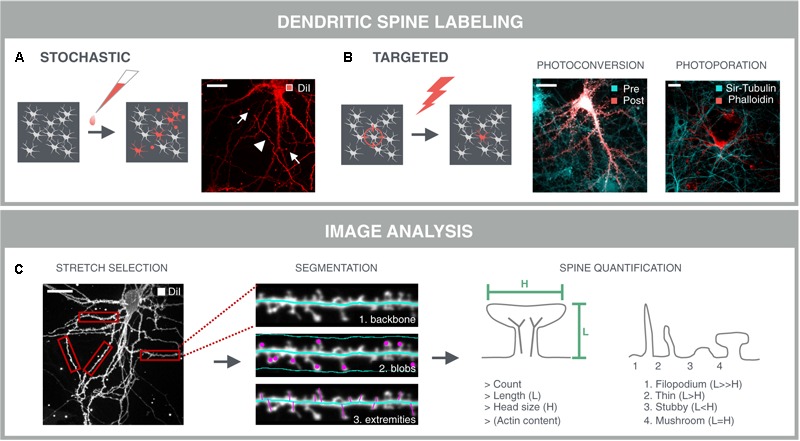
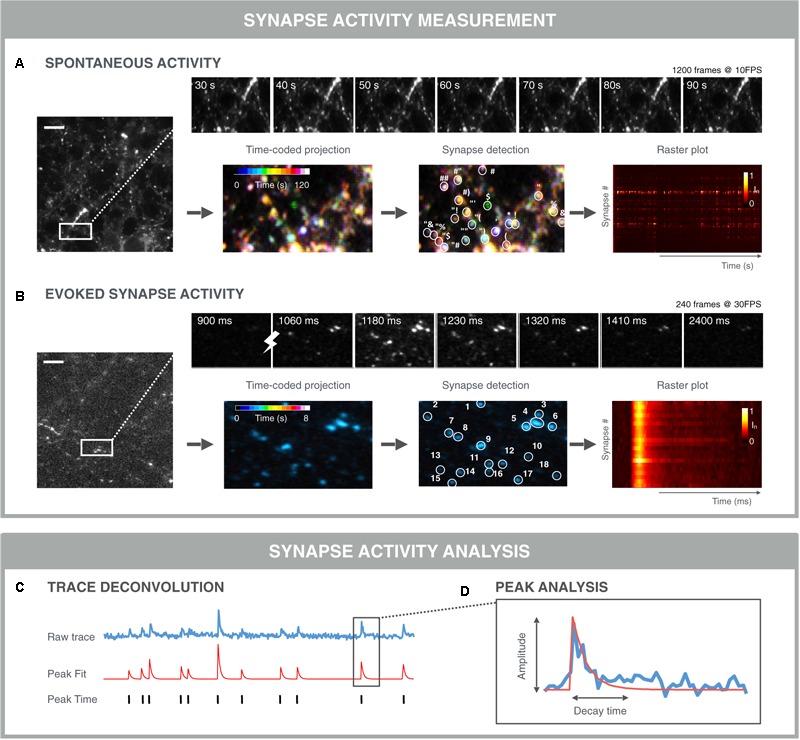
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources