

αv Integrins regulate germinal center B cell responses through noncanonical autophagy
- PMID: 29999501
- PMCID: PMC6118577
- DOI: 10.1172/JCI99597
αv Integrins regulate germinal center B cell responses through noncanonical autophagy
Abstract
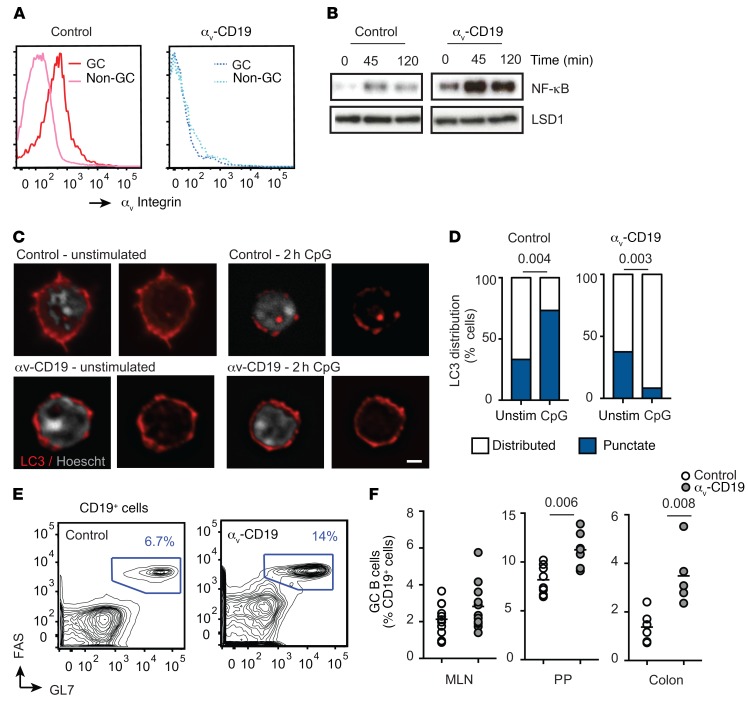
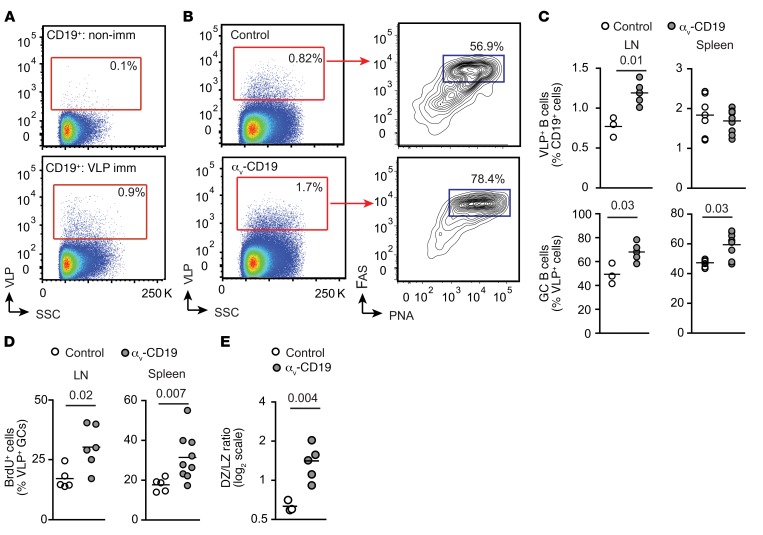
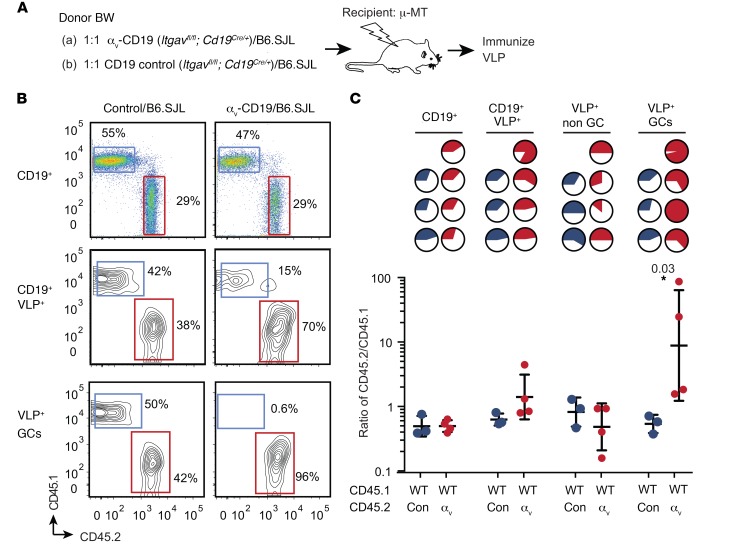
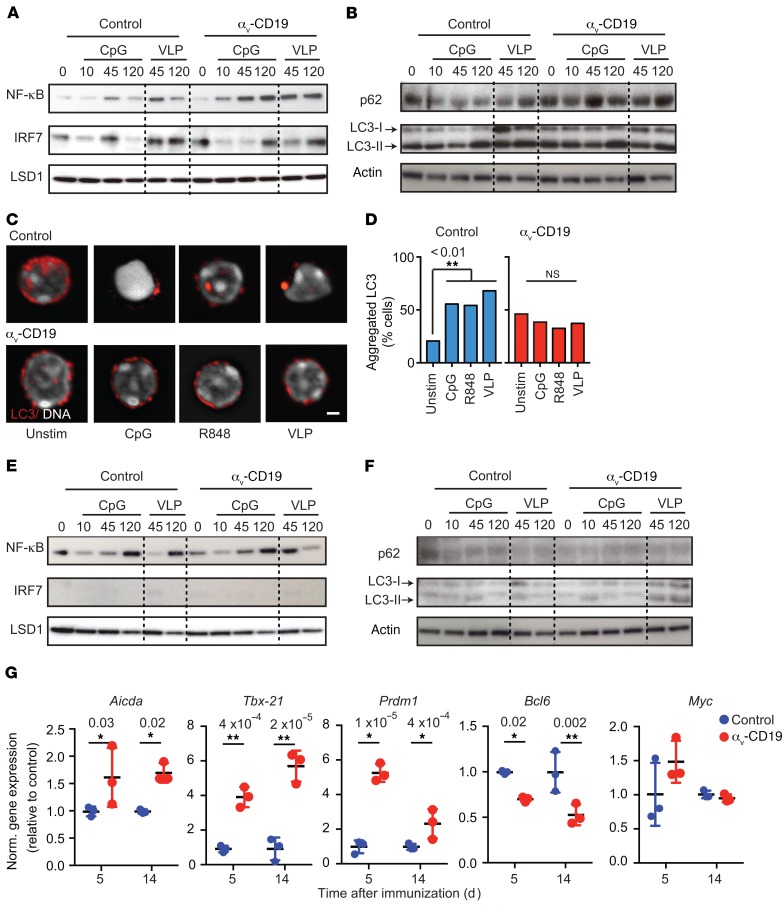
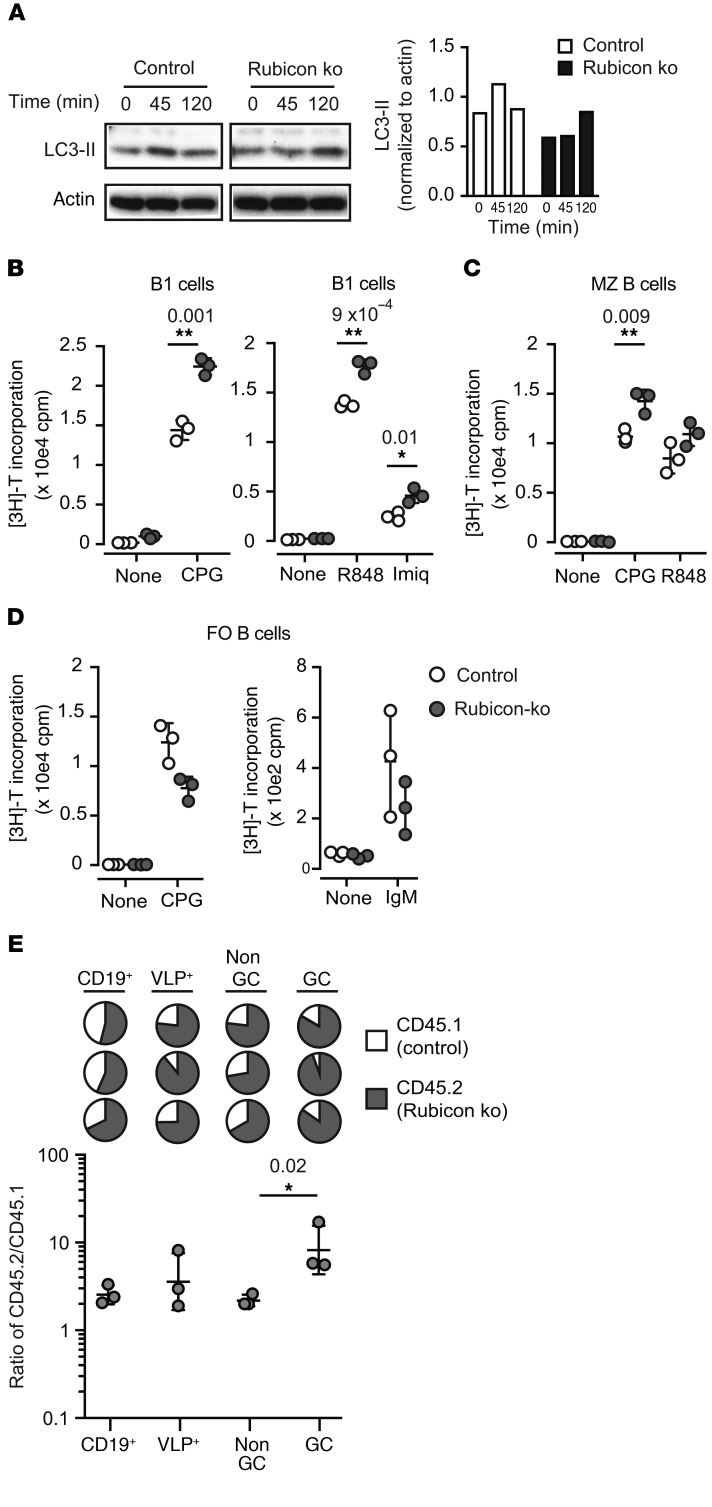
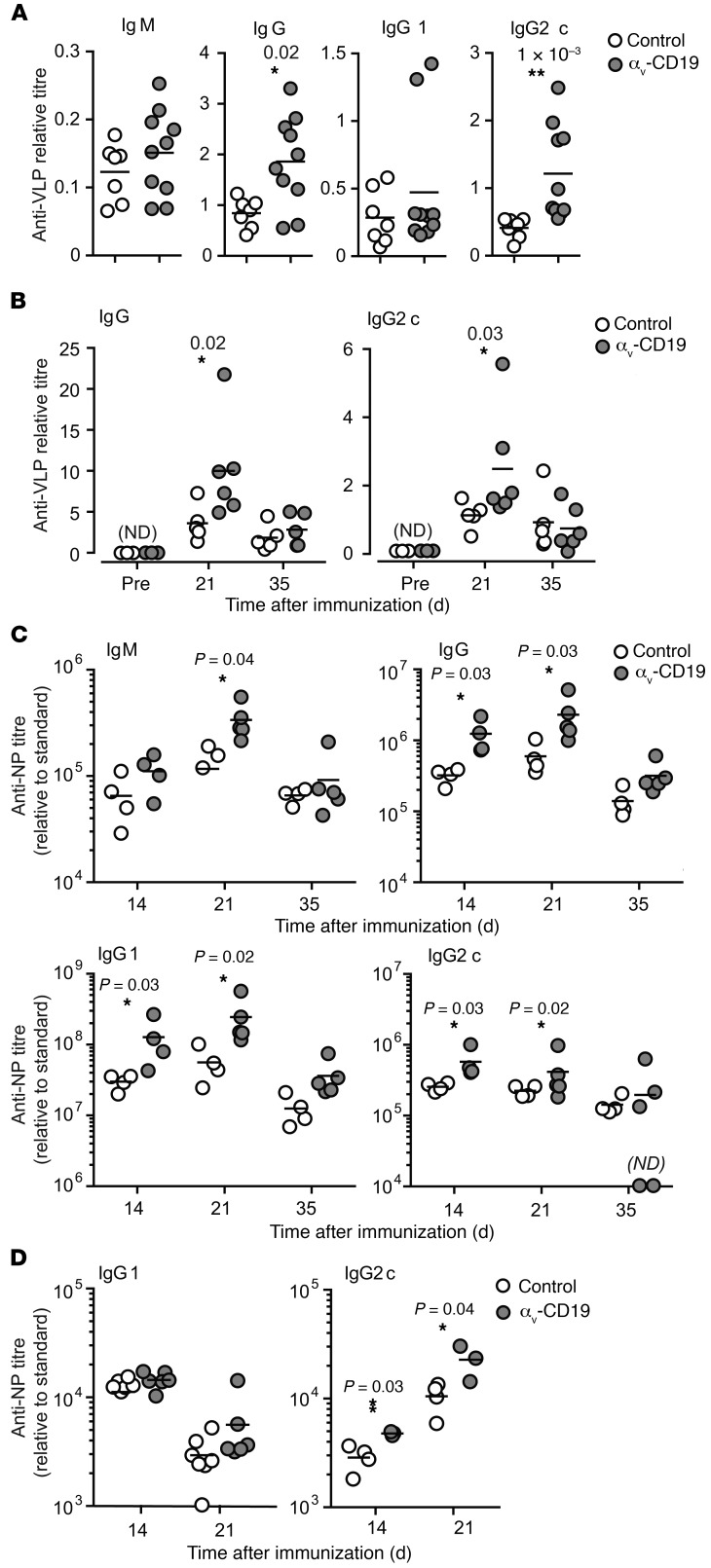
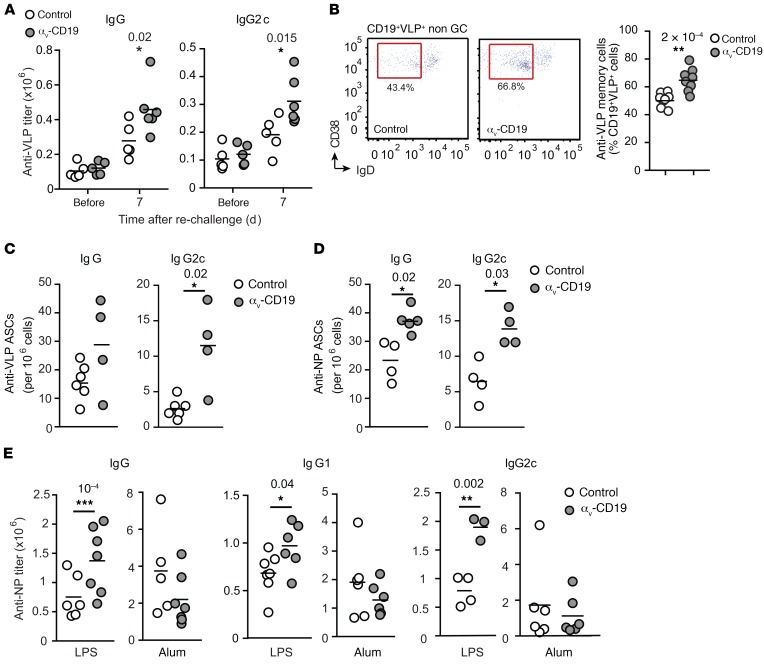
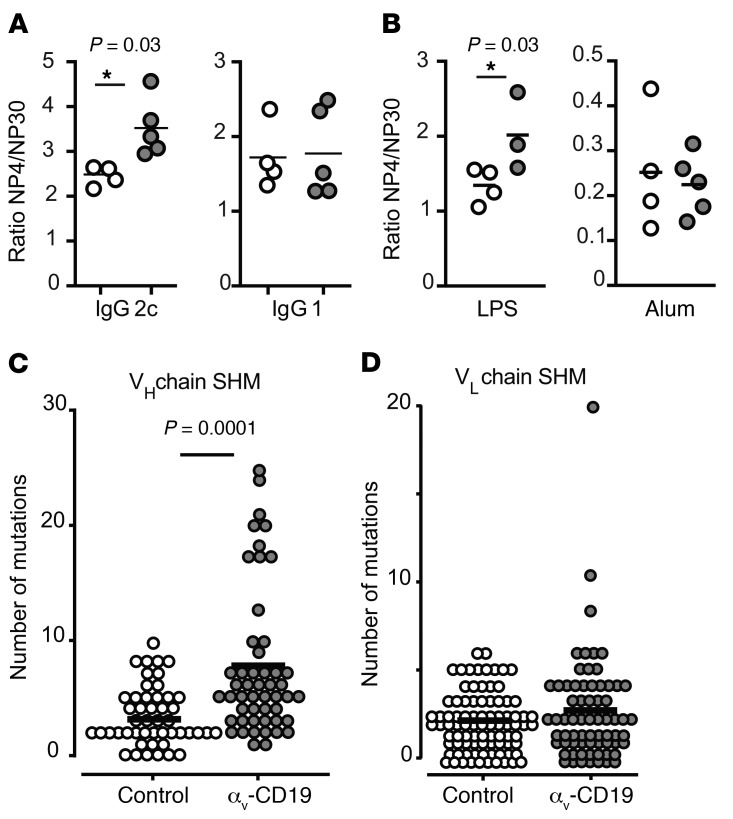
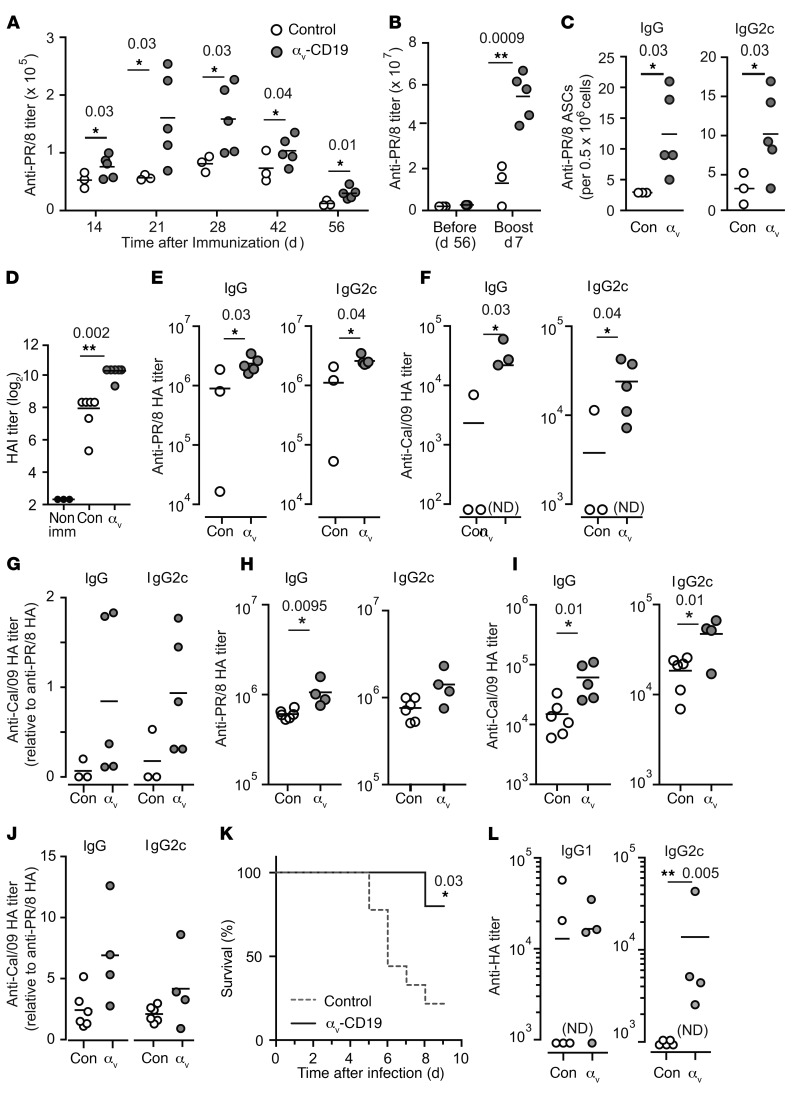
Germinal centers (GCs) are major sites of clonal B cell expansion and generation of long-lived, high-affinity antibody responses to pathogens. Signaling through TLRs on B cells promotes many aspects of GC B cell responses, including affinity maturation, class switching, and differentiation into long-lived memory and plasma cells. A major challenge for effective vaccination is identifying strategies to specifically promote GC B cell responses. Here, we have identified a mechanism of regulation of GC B cell TLR signaling, mediated by αv integrins and noncanonical autophagy. Using B cell-specific αv-KO mice, we show that loss of αv-mediated TLR regulation increased GC B cell expansion, somatic hypermutation, class switching, and generation of long-lived plasma cells after immunization with virus-like particles (VLPs) or antigens associated with TLR ligand adjuvants. Furthermore, targeting αv-mediated regulation increased the magnitude and breadth of antibody responses to influenza virus vaccination. These data therefore identify a mechanism of regulation of GC B cells that can be targeted to enhance antibody responses to vaccination.
Keywords: B cells; Immunology; Innate immunity; Integrins; Vaccines.
Conflict of interest statement
Figures
Comment in
-
Lose appetite, lose control: integrins and noncanonical autophagy regulate germinal center reactions.J Clin Invest. 2018 Aug 31;128(9):3752-3753. doi: 10.1172/JCI122766. Epub 2018 Aug 20. J Clin Invest. 2018. PMID: 30124470 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
