

P2RX7 Purinoceptor as a Therapeutic Target-The Second Coming?
- PMID: 30003075
- PMCID: PMC6032550
- DOI: 10.3389/fchem.2018.00248
P2RX7 Purinoceptor as a Therapeutic Target-The Second Coming?
Abstract
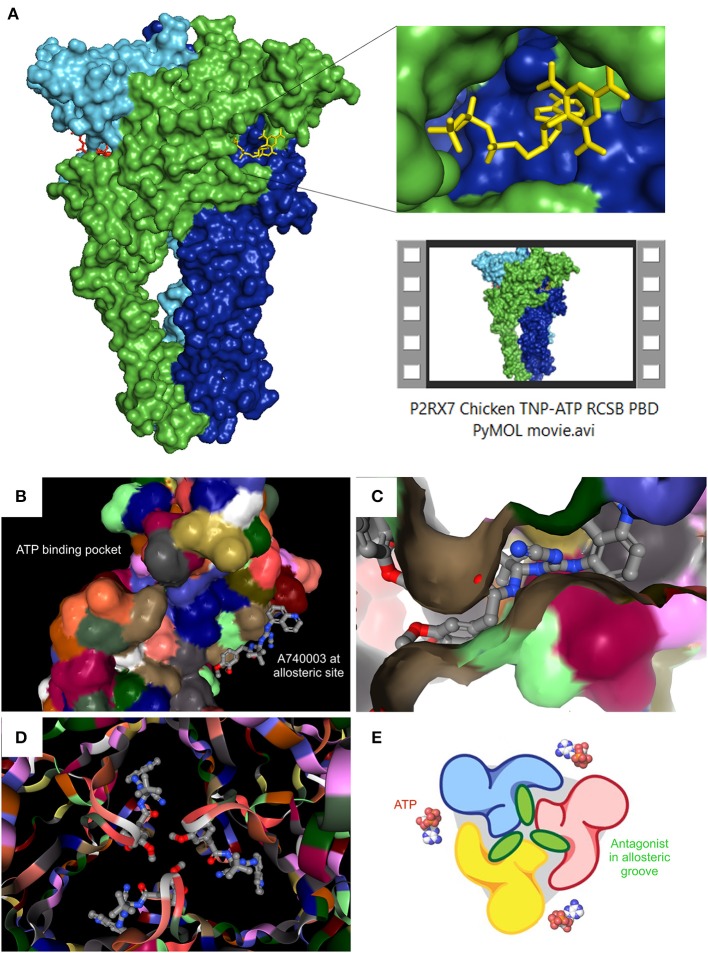
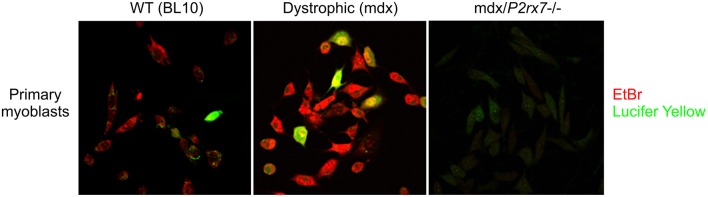
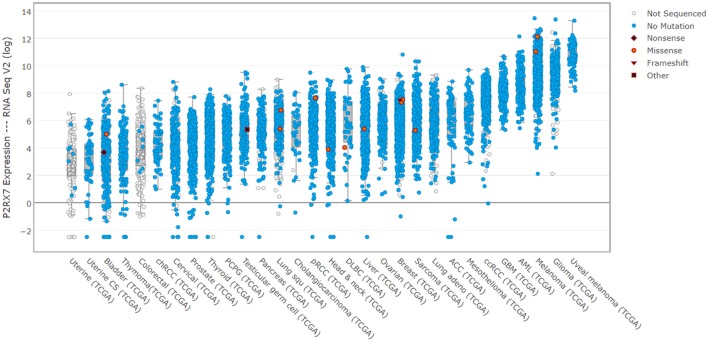
The P2RX7 receptor is a unique member of a family of extracellular ATP (eATP)-gated ion channels expressed in immune cells, where its activation triggers the inflammatory cascade. Therefore, P2RX7 has been long investigated as a target in the treatment of infectious and inflammatory diseases. Subsequently, P2RX7 signaling has been documented in other physiological and pathological processes including pain, CNS and psychiatric disorders and cancer. As a result, a range of P2RX7 antagonists have been developed and trialed. Interestingly, the recent crystallization of mammalian and chicken receptors revealed that most widely-used antagonists may bind a unique allosteric site. The availability of crystal structures allows rational design of improved antagonists and modeling of binding sites of the known or presumed inhibitors. However, several unanswered questions limit the cogent development of P2RX7 therapies. Firstly, this receptor functions as an ion channel, but its chronic stimulation by high eATP causes opening of the non-selective large pore (LP), which can trigger cell death. Not only the molecular mechanism of LP opening is still not fully understood but its function(s) are also unclear. Furthermore, how can tumor cells take advantage of P2RX7 for growth and spread and yet survive overexpression of potentially cytotoxic LP in the eATP-rich environment? The recent discovery of the feedback loop, wherein the LP-evoked release of active MMP-2 triggers the receptor cleavage, provided one explanation. Another mechanism might be that of cancer cells expressing a structurally altered P2RX7 receptor, devoid of the LP function. Exploiting such mechanisms should lead to the development of new, less toxic anticancer treatments. Notably, targeted inhibition of P2RX7 is crucial as its global blockade reduces the immune and inflammatory responses, which have important anti-tumor effects in some types of malignancies. Therefore, another novel approach is the synthesis of tissue/cell specific P2RX7 antagonists. Progress has been aided by the development of p2rx7 knockout mice and new conditional knock-in and knock-out models are being created. In this review, we seek to summarize the recent advances in our understanding of molecular mechanisms of receptor activation and inhibition, which cause its re-emergence as an important therapeutic target. We also highlight the key difficulties affecting this development.
Keywords: ATP; DMD; P2RX7 KO mouse; P2X7; cancer; inflammation; large pore.
Figures
Similar articles
-
A novel mechanism of autophagic cell death in dystrophic muscle regulated by P2RX7 receptor large-pore formation and HSP90.Autophagy. 2015;11(1):113-30. doi: 10.4161/15548627.2014.994402. Autophagy. 2015. PMID: 25700737 Free PMC article.
-
P2RX7 purinoceptor: a therapeutic target for ameliorating the symptoms of duchenne muscular dystrophy.PLoS Med. 2015 Oct 13;12(10):e1001888. doi: 10.1371/journal.pmed.1001888. eCollection 2015 Oct. PLoS Med. 2015. PMID: 26461208 Free PMC article.
-
Zidovudine ameliorates pathology in the mouse model of Duchenne muscular dystrophy via P2RX7 purinoceptor antagonism.Acta Neuropathol Commun. 2018 Apr 11;6(1):27. doi: 10.1186/s40478-018-0530-4. Acta Neuropathol Commun. 2018. PMID: 29642926 Free PMC article.
-
P2X7 receptors: channels, pores and more.CNS Neurol Disord Drug Targets. 2012 Sep;11(6):705-21. doi: 10.2174/187152712803581137. CNS Neurol Disord Drug Targets. 2012. PMID: 22963440 Review.
-
P2X7 purinoceptor as a therapeutic target in muscular dystrophies.Curr Opin Pharmacol. 2019 Aug;47:40-45. doi: 10.1016/j.coph.2019.02.003. Epub 2019 Mar 20. Curr Opin Pharmacol. 2019. PMID: 30901735 Review.
Cited by
-
P2X7 Receptors Amplify CNS Damage in Neurodegenerative Diseases.Int J Mol Sci. 2020 Aug 20;21(17):5996. doi: 10.3390/ijms21175996. Int J Mol Sci. 2020. PMID: 32825423 Free PMC article. Review.
-
The Role of Purinergic P2X7 Receptor in Inflammation and Cancer: Novel Molecular Insights and Clinical Applications.Cancers (Basel). 2022 Feb 22;14(5):1116. doi: 10.3390/cancers14051116. Cancers (Basel). 2022. PMID: 35267424 Free PMC article. Review.
-
Cancer Metabostemness and Metabolic Reprogramming via P2X7 Receptor.Cells. 2021 Jul 14;10(7):1782. doi: 10.3390/cells10071782. Cells. 2021. PMID: 34359950 Free PMC article. Review.
-
How Structural Biology Has Directly Impacted Our Understanding of P2X Receptor Function and Gating.Methods Mol Biol. 2022;2510:1-29. doi: 10.1007/978-1-0716-2384-8_1. Methods Mol Biol. 2022. PMID: 35776317
-
Clemastine fumarate accelerates accumulation of disability in progressive multiple sclerosis by enhancing pyroptosis.medRxiv [Preprint]. 2024 Apr 10:2024.04.09.24305506. doi: 10.1101/2024.04.09.24305506. medRxiv. 2024. Update in: J Clin Invest. 2025 May 15;135(10):e183941. doi: 10.1172/JCI183941. PMID: 39802756 Free PMC article. Updated. Preprint.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous