

Central sensitization of the spino-parabrachial-amygdala pathway that outlasts a brief nociceptive stimulus
- PMID: 30004124
- PMCID: PMC6138295
- DOI: 10.1113/JP273976
Central sensitization of the spino-parabrachial-amygdala pathway that outlasts a brief nociceptive stimulus
Abstract
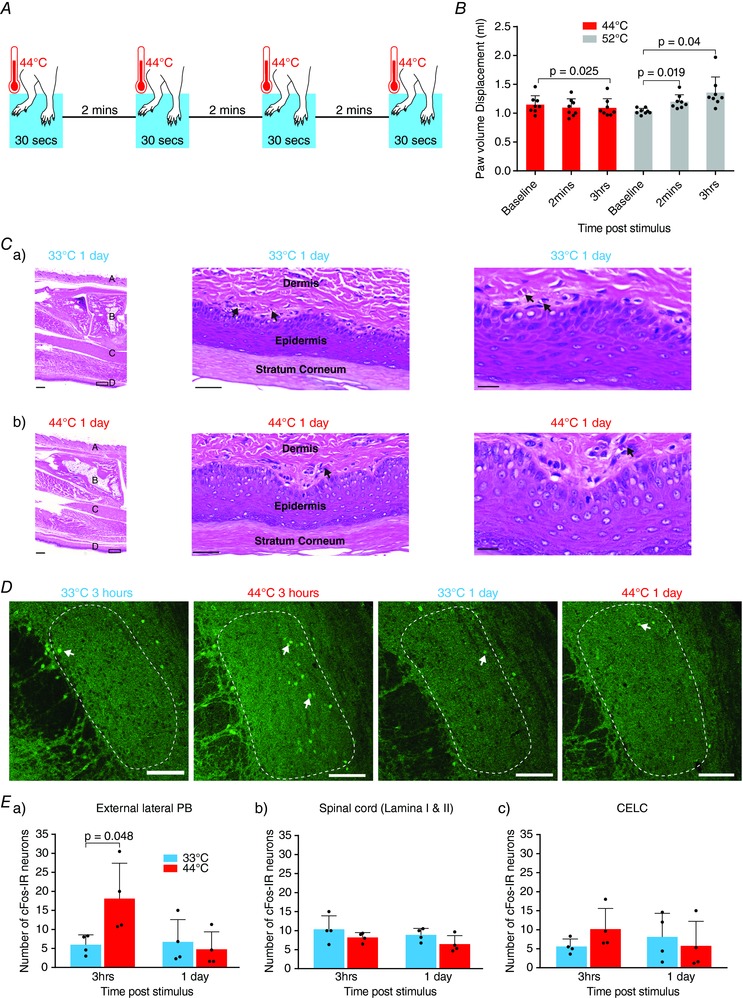
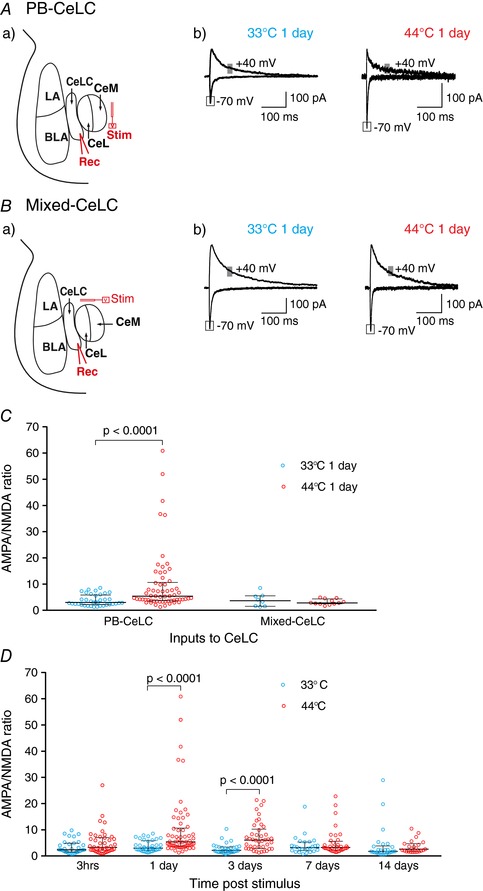
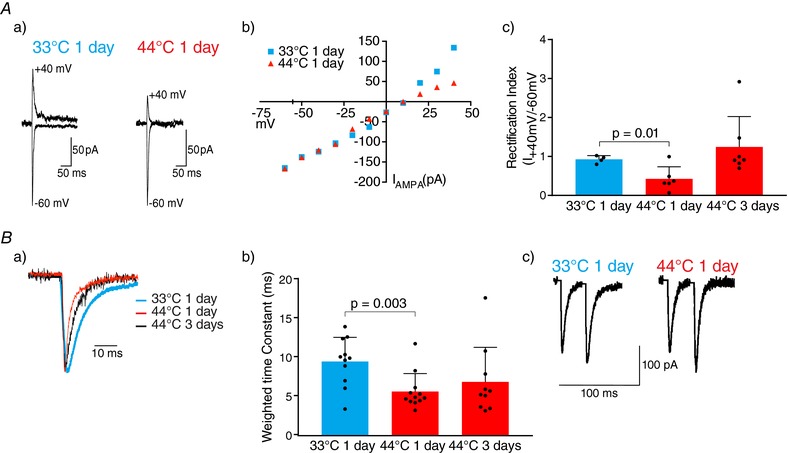
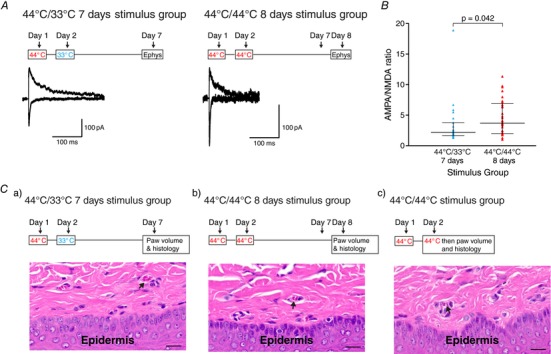
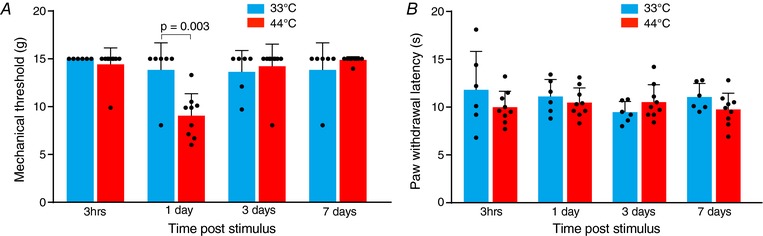
Key points: Chronic pain is disabling because sufferers form negative associations between pain and activities, such as work, leading to the sufferer limiting these activities. Pain information arriving in the amygdala is responsible for forming these associations and contributes to us feeling bad when we are in pain. Ongoing injuries enhance the delivery of pain information to the amygdala. If we want to understand why chronic pain can continue without ongoing injury, it is important to know whether this facilitation continues once the injury has healed. In the present study, we show that a 2 min noxious heat stimulus, without ongoing injury, is able to enhance delivery of pain information to the amygdala for 3 days. If the noxious heat stimulus is repeated, this enhancement persists even longer. These changes may prime this information pathway so that subsequent injuries may feel even worse and the associative learning that results in pain-related avoidance may be promoted.
Abstract: Pain is an important defence against dangers in our environment; however, some clinical conditions produce pain that outlasts this useful role and persists even after the injury has healed. The experience of pain consists of somatosensory elements of intensity and location, negative emotional/aversive feelings and subsequent restrictions on lifestyle as a result of a learned association between certain activities and pain. The amygdala contributes negative emotional value to nociceptive sensory information and forms the association between an aversive response and the environment in which it occurs. It is able to form this association because it receives nociceptive information via the spino-parabrachio-amygdaloid pathway and polymodal sensory information via cortical and thalamic inputs. Synaptic plasticity occurs at the parabrachial-amygdala synapse and other brain regions in chronic pain conditions with ongoing injury; however, very little is known about how plasticity occurs in conditions with no ongoing injury. Using immunohistochemistry, electrophysiology and behavioural assays, we show that a brief nociceptive stimulus with no ongoing injury is able to produce long-lasting synaptic plasticity at the rat parabrachial-amygdala synapse. We show that this plasticity is caused by an increase in postsynaptic AMPA receptors with a transient change in the AMPA receptor subunit, similar to long-term potentiation. Furthermore, this synaptic potentiation primes the synapse so that a subsequent noxious stimulus causes prolonged potentiation of the nociceptive information flow into the amygdala. As a result, a second injury could have an increased negative emotional value and promote associative learning that results in pain-related avoidance.
Keywords: Synaptic plasticity; ampa receptor; amygdala; pain; parabrachial nucleus.
© 2018 The Authors. The Journal of Physiology © 2018 The Physiological Society.
Figures
Similar articles
-
Predominant synaptic potentiation and activation in the right central amygdala are independent of bilateral parabrachial activation in the hemilateral trigeminal inflammatory pain model of rats.Mol Pain. 2018 Jan-Dec;14:1744806918807102. doi: 10.1177/1744806918807102. Epub 2018 Oct 1. Mol Pain. 2018. PMID: 30270724 Free PMC article.
-
Synaptic potentiation in the nociceptive amygdala following fear learning in mice.Mol Brain. 2013 Mar 1;6:11. doi: 10.1186/1756-6606-6-11. Mol Brain. 2013. PMID: 23452928 Free PMC article.
-
Processing of nociceptive mechanical and thermal information in central amygdala neurons with knee-joint input.J Neurophysiol. 2002 Jan;87(1):103-12. doi: 10.1152/jn.00264.2001. J Neurophysiol. 2002. PMID: 11784733
-
Left and right hemispheric lateralization of the amygdala in pain.Prog Neurobiol. 2021 Jan;196:101891. doi: 10.1016/j.pneurobio.2020.101891. Epub 2020 Jul 28. Prog Neurobiol. 2021. PMID: 32730859 Free PMC article. Review.
-
Short-term synaptic plasticity in the nociceptive thalamic-anterior cingulate pathway.Mol Pain. 2009 Sep 4;5:51. doi: 10.1186/1744-8069-5-51. Mol Pain. 2009. PMID: 19732417 Free PMC article. Review.
Cited by
-
Structural and temporal dynamics analysis of neural circuit from 2002 to 2022: A bibliometric analysis.Heliyon. 2024 Jan 14;10(2):e24649. doi: 10.1016/j.heliyon.2024.e24649. eCollection 2024 Jan 30. Heliyon. 2024. PMID: 38298625 Free PMC article. Review.
-
Dysfunction of Small-Conductance Ca2+-Activated Potassium (SK) Channels Drives Amygdala Hyperexcitability and Neuropathic Pain Behaviors: Involvement of Epigenetic Mechanisms.Cells. 2024 Jun 18;13(12):1055. doi: 10.3390/cells13121055. Cells. 2024. PMID: 38920682 Free PMC article.
-
Tonic pain alters functional connectivity of the descending pain modulatory network involving amygdala, periaqueductal gray, parabrachial nucleus and anterior cingulate cortex.Neuroimage. 2022 Aug 1;256:119278. doi: 10.1016/j.neuroimage.2022.119278. Epub 2022 May 9. Neuroimage. 2022. PMID: 35523367 Free PMC article.
-
Danger and distress: Parabrachial-extended amygdala circuits.Neuropharmacology. 2021 Oct 15;198:108757. doi: 10.1016/j.neuropharm.2021.108757. Epub 2021 Aug 27. Neuropharmacology. 2021. PMID: 34461068 Free PMC article. Review.
-
Dopamine receptor D2 regulates GLUA1-containing AMPA receptor trafficking and central sensitization through the PI3K signaling pathway in a male rat model of chronic migraine.J Headache Pain. 2022 Aug 10;23(1):98. doi: 10.1186/s10194-022-01469-x. J Headache Pain. 2022. PMID: 35948867 Free PMC article.
References
-
- Abraham WC. (2008). Metaplasticity: tuning synapses and networks for plasticity. Nat Rev Neurosci 9, 387. - PubMed
-
- Adriaensen H, Gybels J, Handwerker HO & Van Hees J. (1984). Suppression of C‐fibre discharges upon repeated heat stimulation may explain characteristics of concomitant pain sensations. Brain Res 302, 203–211. - PubMed
-
- Ansah OB, Bourbia N, Goncalves L, Almeida A & Pertovaara A. (2010). Influence of amygdaloid glutamatergic receptors on sensory and emotional pain‐related behavior in the neuropathic rat. Behav Brain Res 209, 174–178. - PubMed
-
- Archibald K, Perry MJ, Molnar E & Henley JM. (1998). Surface expression and metabolic half‐life of AMPA receptors in cultured rat cerebellar granule cells. Neuropharmacology 37, 1345–1353. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials