

SLC6A14, an amino acid transporter, modifies the primary CF defect in fluid secretion
- PMID: 30004386
- PMCID: PMC6054531
- DOI: 10.7554/eLife.37963
SLC6A14, an amino acid transporter, modifies the primary CF defect in fluid secretion
Abstract
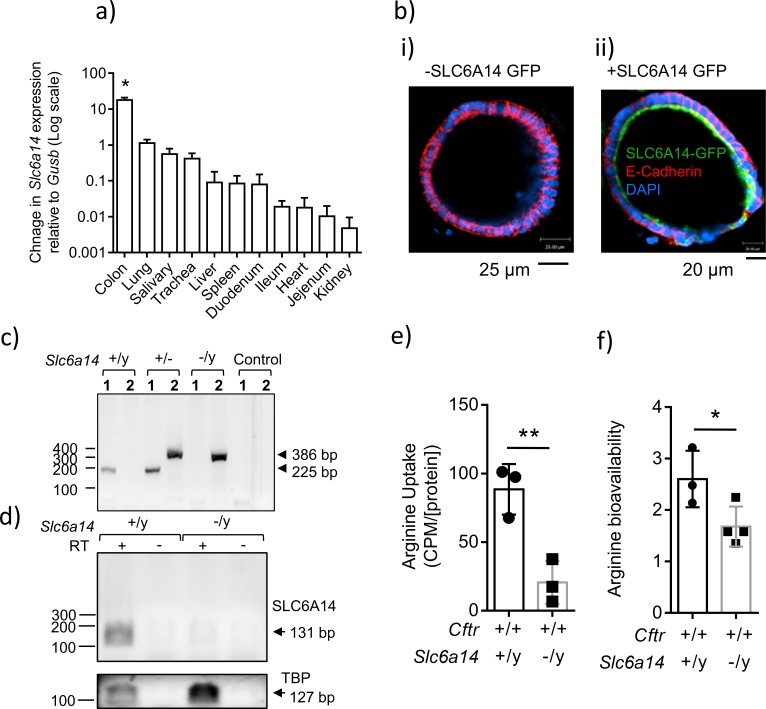
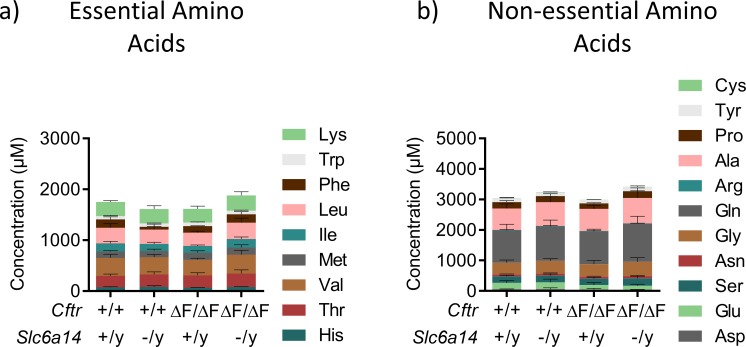
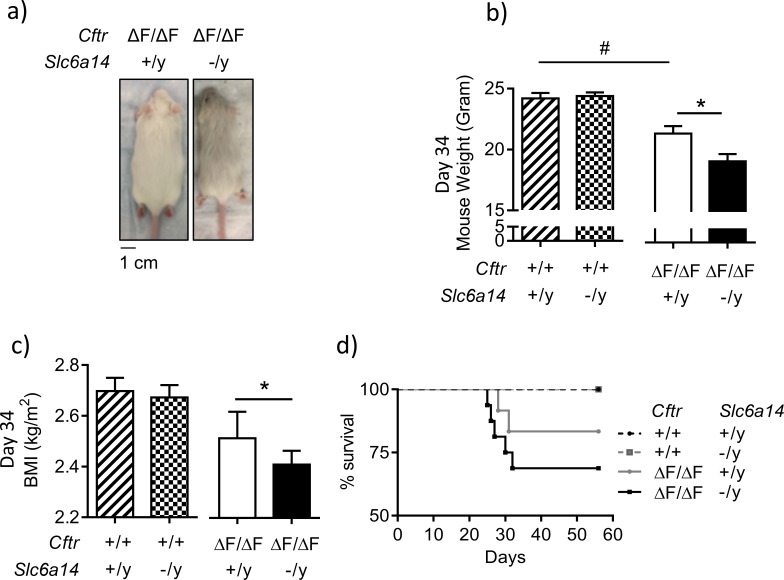
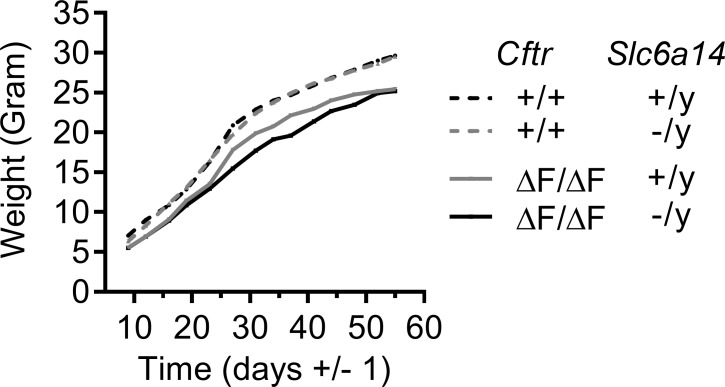
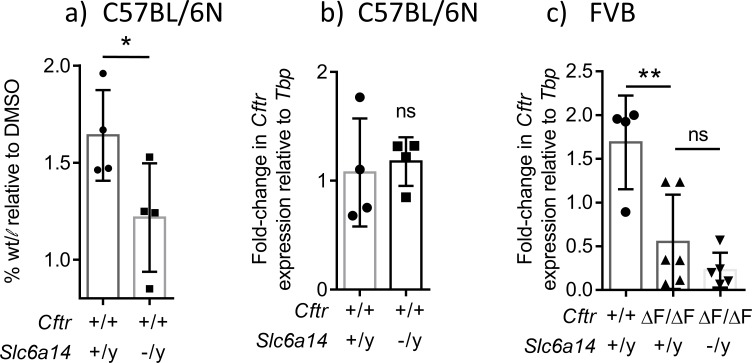
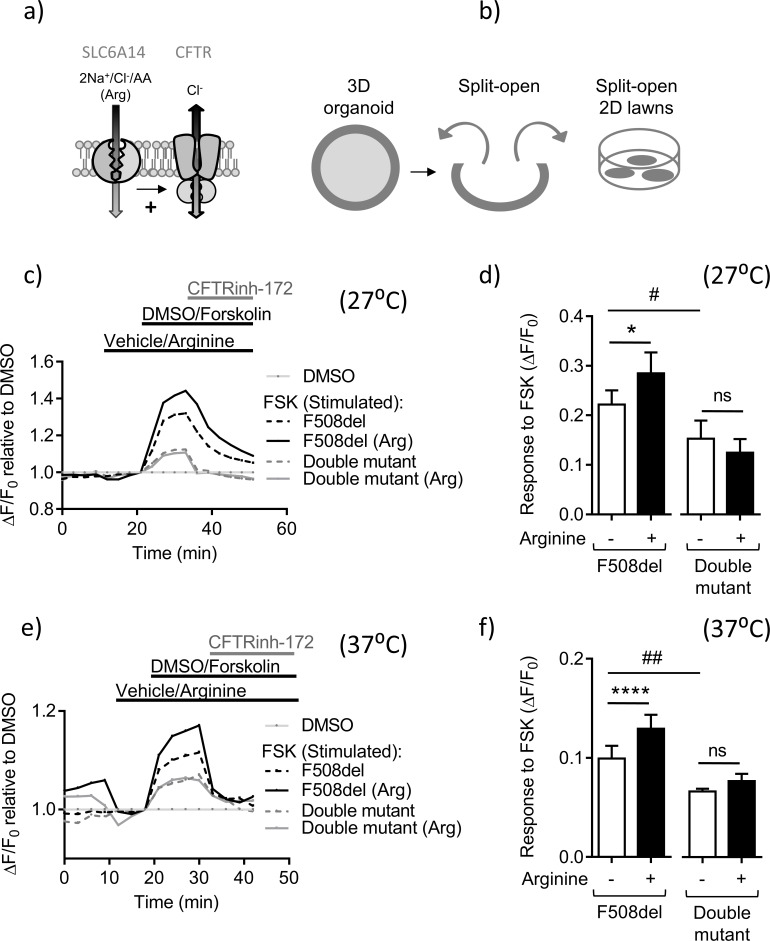
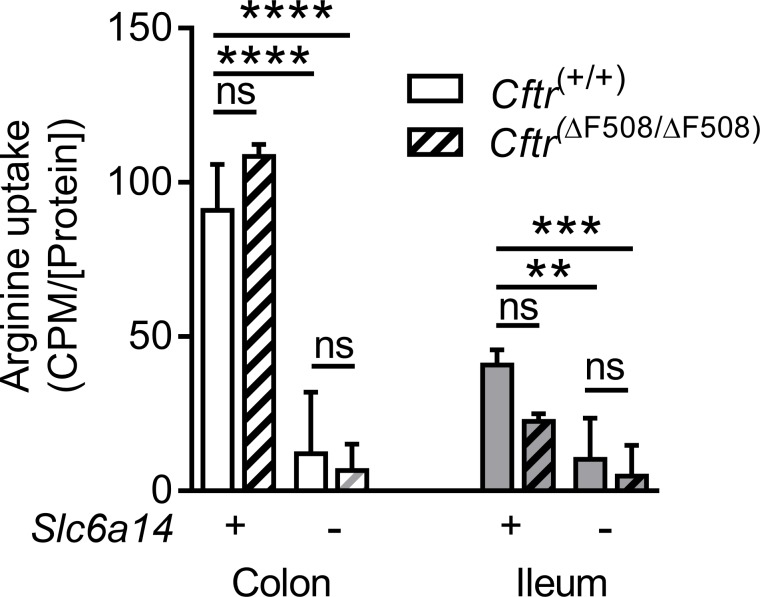
The severity of intestinal disease associated with Cystic Fibrosis (CF) is variable in the patient population and this variability is partially conferred by the influence of modifier genes. Genome-wide association studies have identified SLC6A14, an electrogenic amino acid transporter, as a genetic modifier of CF-associated meconium ileus. The purpose of the current work was to determine the biological role of Slc6a14, by disrupting its expression in CF mice bearing the major mutation, F508del. We found that disruption of Slc6a14 worsened the intestinal fluid secretion defect, characteristic of these mice. In vitro studies of mouse intestinal organoids revealed that exacerbation of the primary defect was associated with reduced arginine uptake across the apical membrane, with aberrant nitric oxide and cyclic GMP-mediated regulation of the major CF-causing mutant protein. Together, these studies highlight the role of this apical transporter in modifying cellular nitric oxide levels, residual function of the major CF mutant and potentially, its promise as a therapeutic target.
Keywords: F508del-CFTR; amino acid transporter; colonic cell line; human biology; medicine; mouse; nitric oxide signaling; organoids.
© 2018, Ahmadi et al.
Conflict of interest statement
SA, SX, YW, MD, RK, CL, FL, KD, JR, CB No competing interests declared
Figures
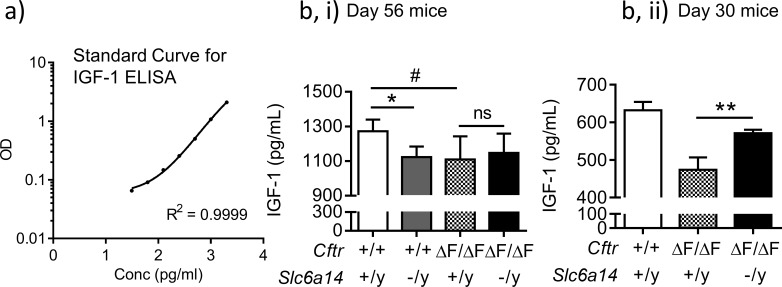
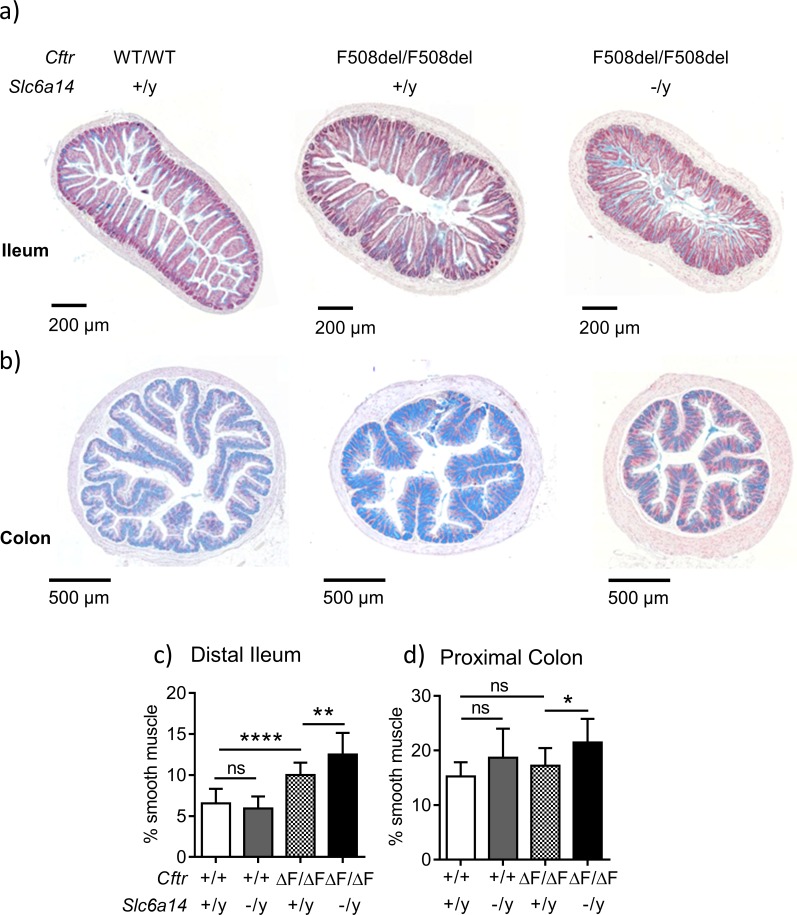
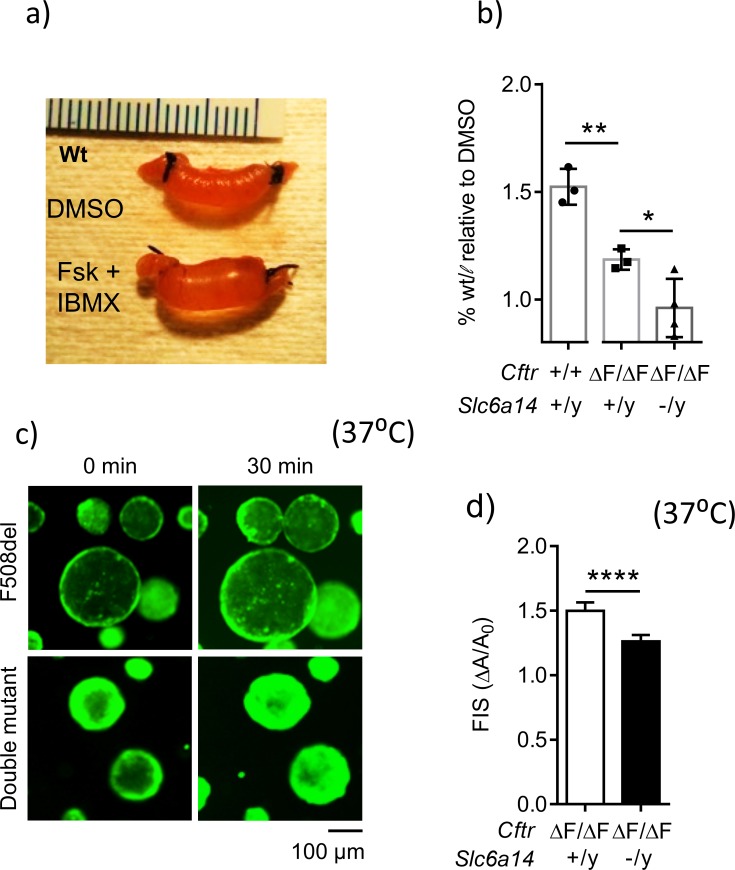
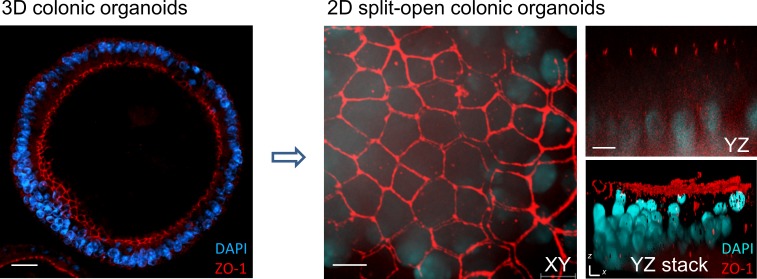
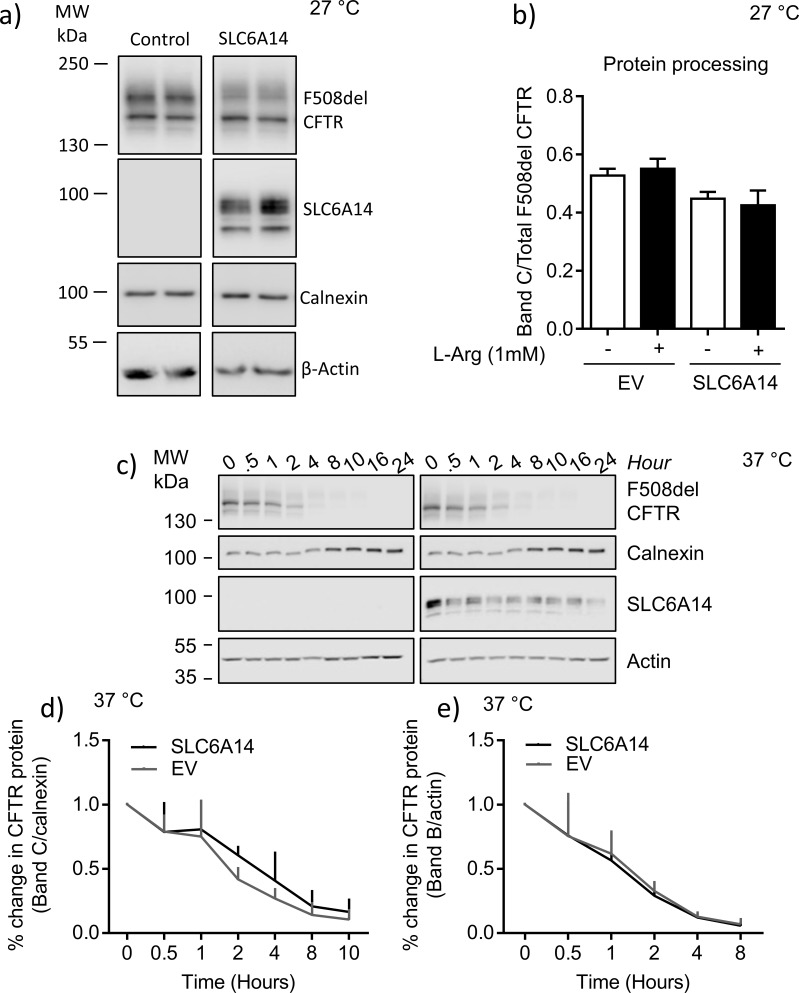
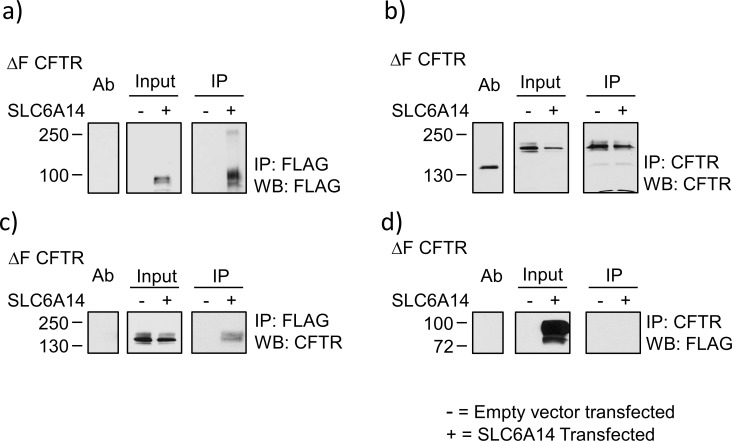
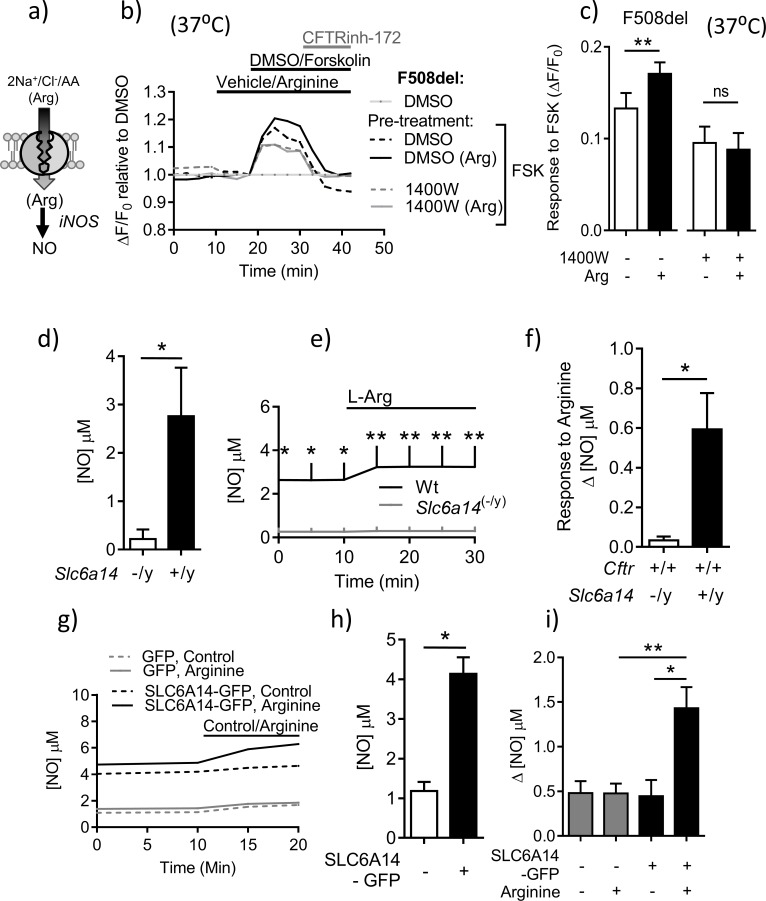
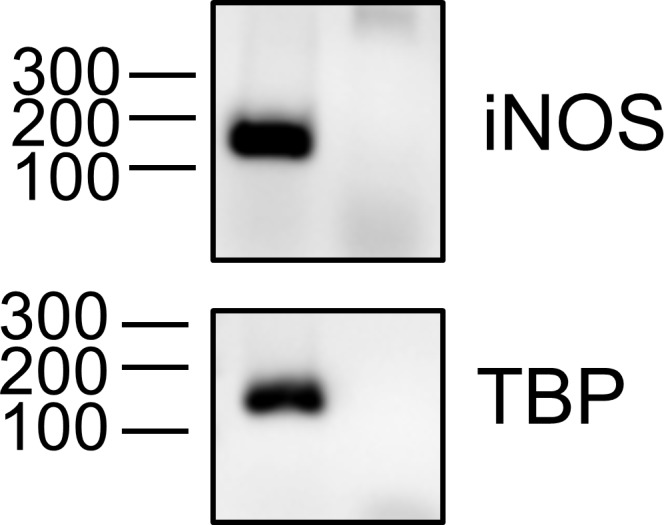
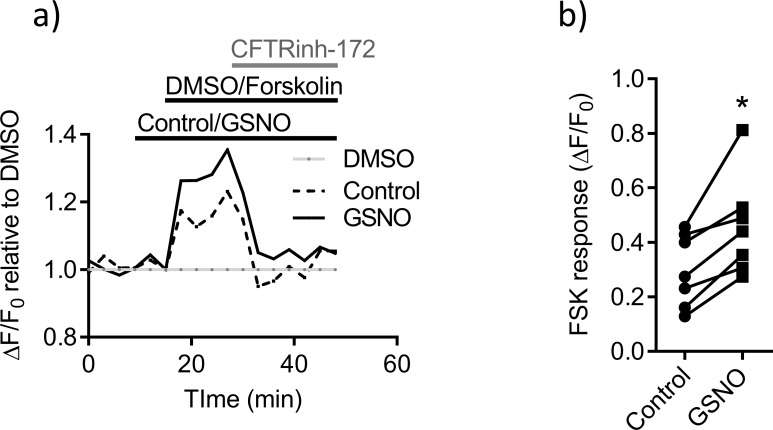
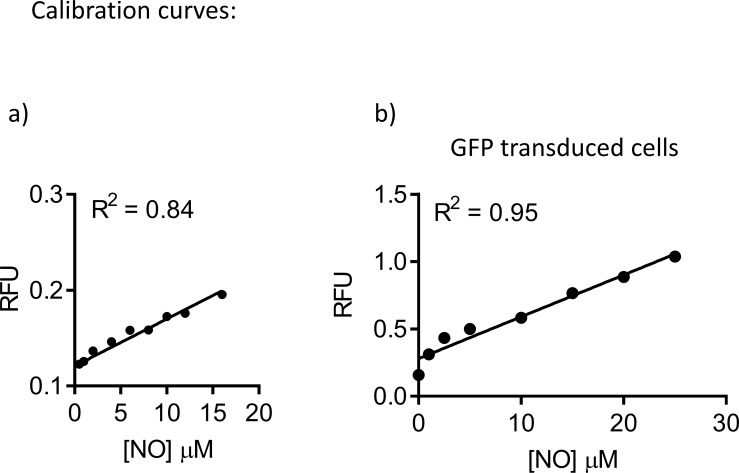
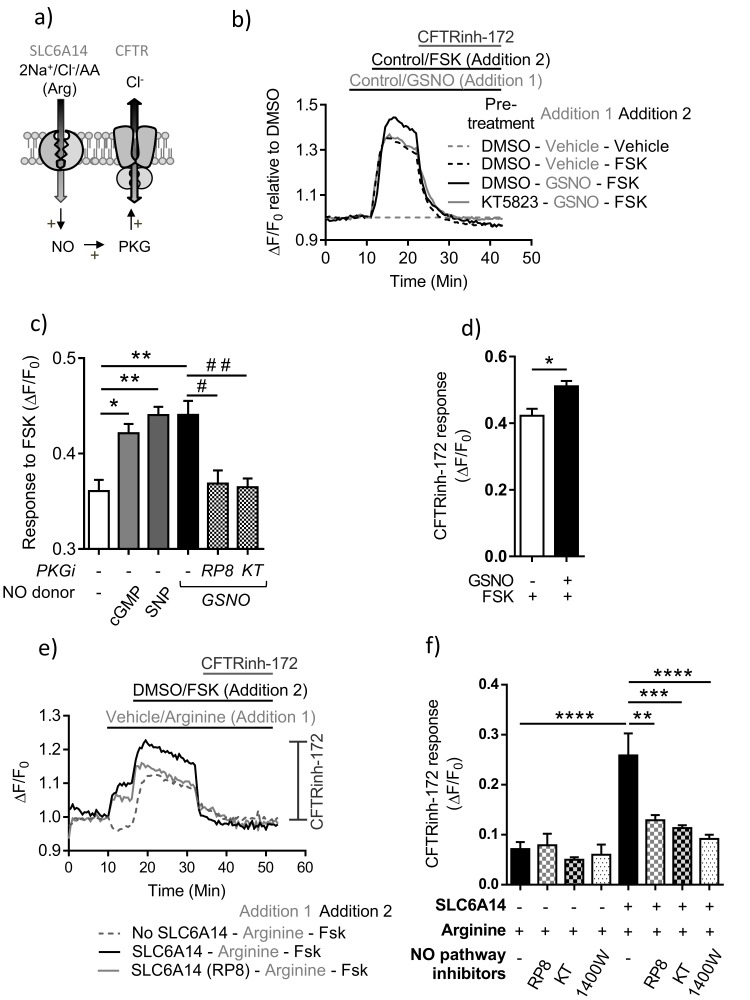
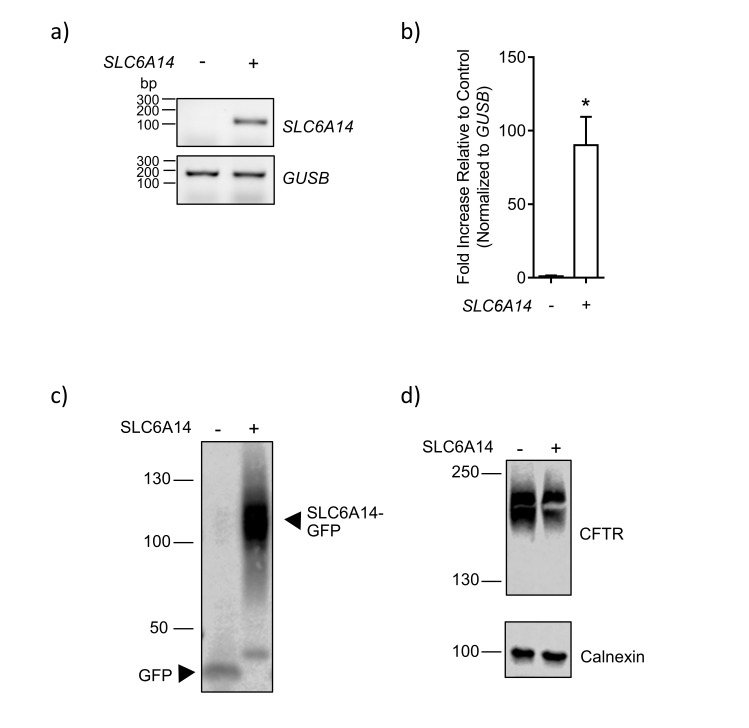
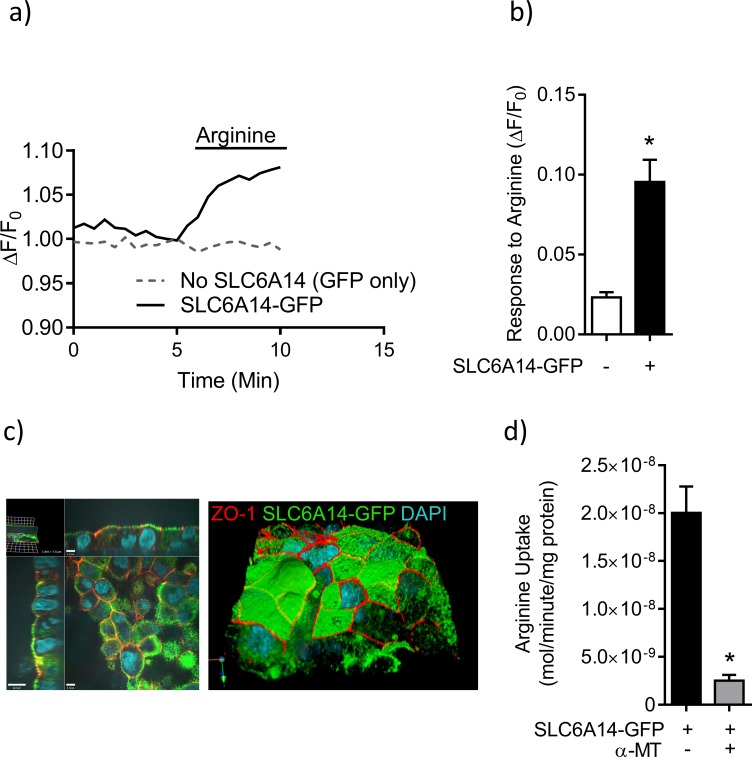
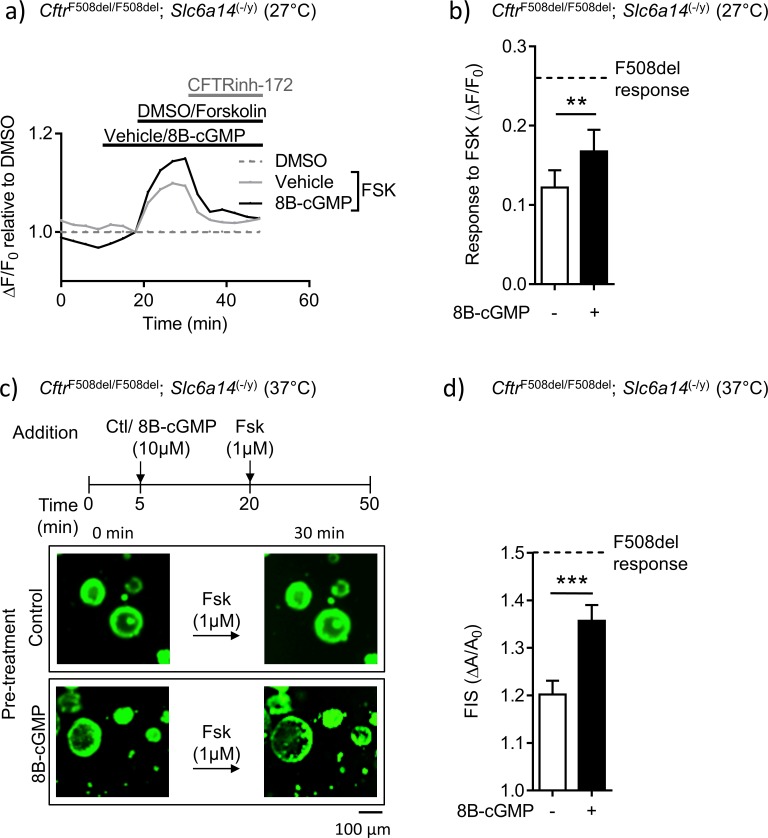
References
-
- Ahmadi S, Bozoky Z, Di Paola M, Xia S, Li C, Wong AP, Wellhauser L, Molinski SV, Ip W, Ouyang H, Avolio J, Forman-Kay JD, Ratjen F, Hirota JA, Rommens J, Rossant J, Gonska T, Moraes TJ, Bear CE. Phenotypic profiling of CFTR modulators in patient-derived respiratory epithelia. npj Genomic Medicine. 2017;2:12. doi: 10.1038/s41525-017-0015-6. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
