

Calcium-activated chloride channels clamp odor-evoked spike activity in olfactory receptor neurons
- PMID: 30006552
- PMCID: PMC6045664
- DOI: 10.1038/s41598-018-28855-3
Calcium-activated chloride channels clamp odor-evoked spike activity in olfactory receptor neurons
Abstract
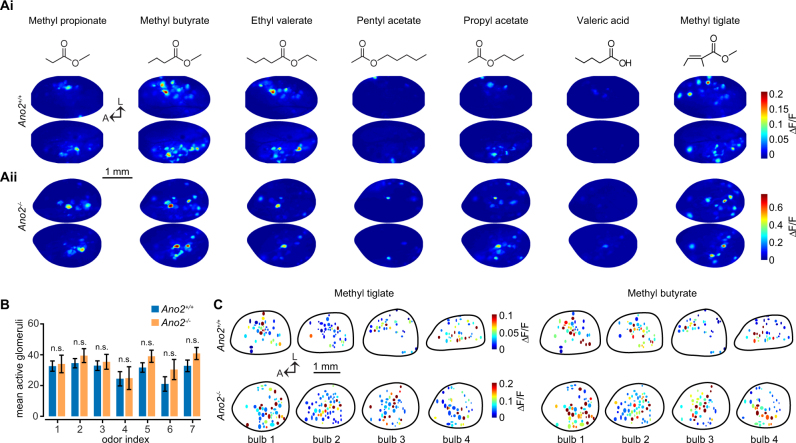
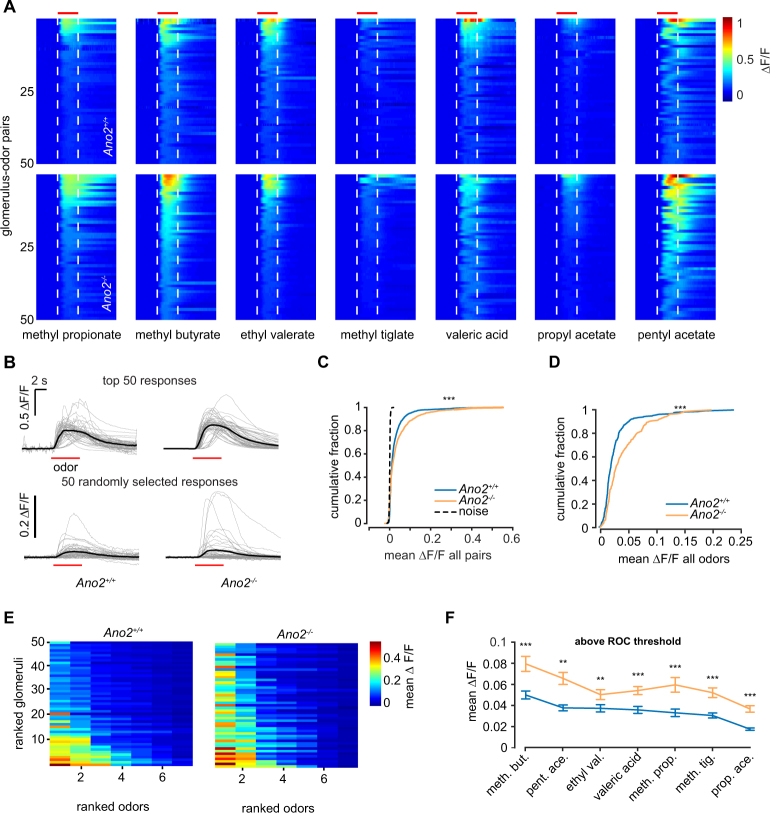
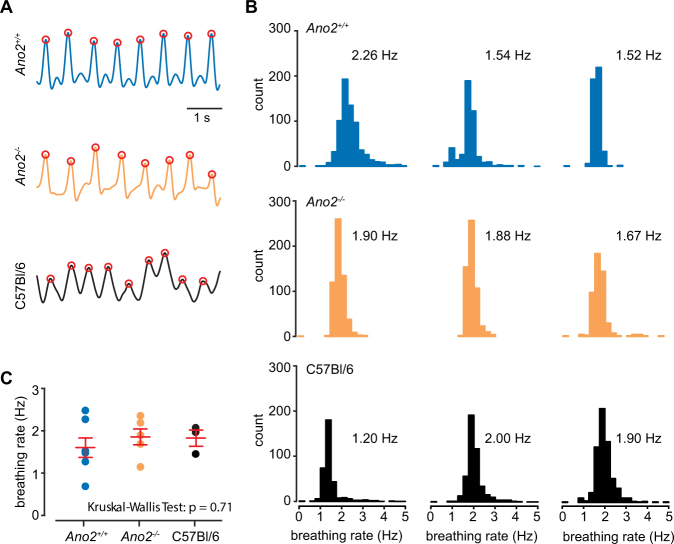
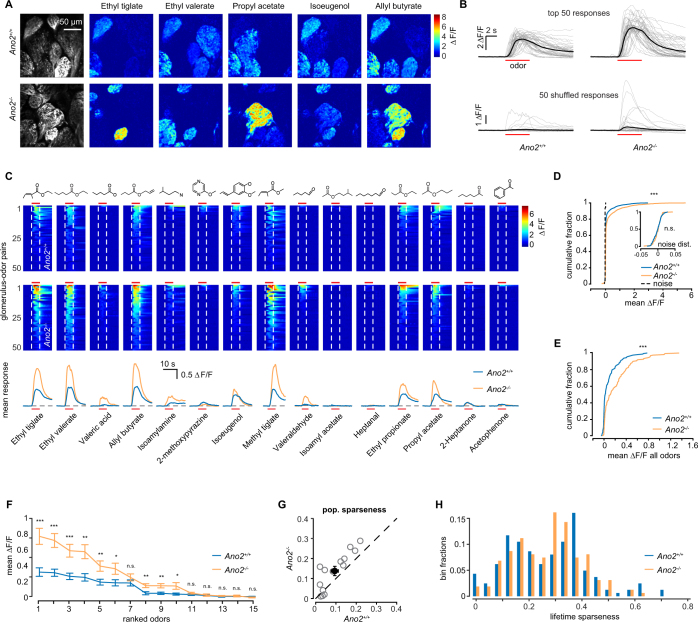
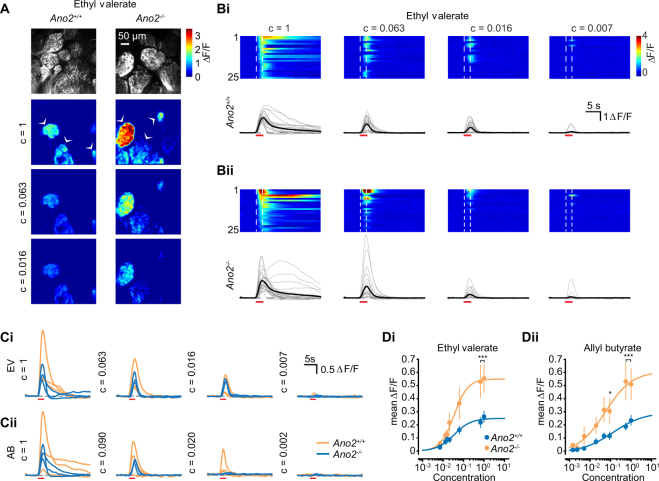
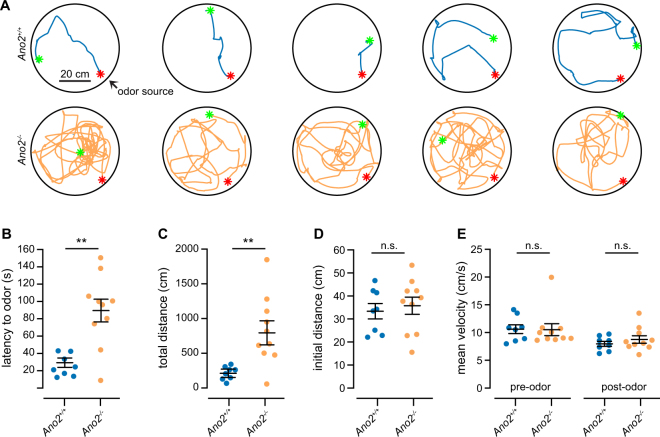
The calcium-activated chloride channel anoctamin-2 (Ano2) is thought to amplify transduction currents in olfactory receptor neurons (ORNs), a hypothesis supported by previous studies in dissociated neurons from Ano2-/- mice. Paradoxically, despite a reduction in transduction currents in Ano2-/- ORNs, their spike output for odor stimuli may be higher. We examined the role of Ano2 in ORNs in their native environment in freely breathing mice by imaging activity in ORN axons as they arrive in the olfactory bulb glomeruli. Odor-evoked responses in ORN axons of Ano2-/- animals were consistently larger for a variety of odorants and concentrations. In an open arena, Ano2-/- animals took longer to approach a localized odor source than Ano2+/+ animals, revealing clear olfactory behavioral deficits. Our studies provide the first in vivo evidence toward an alternative or additional role for Ano2 in the olfactory transduction cascade, where it may serve as a feedback mechanism to clamp ORN spike output.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
