

Molecular mechanisms of force production in clathrin-mediated endocytosis
- PMID: 30006986
- PMCID: PMC6231980
- DOI: 10.1002/1873-3468.13192
Molecular mechanisms of force production in clathrin-mediated endocytosis
Abstract
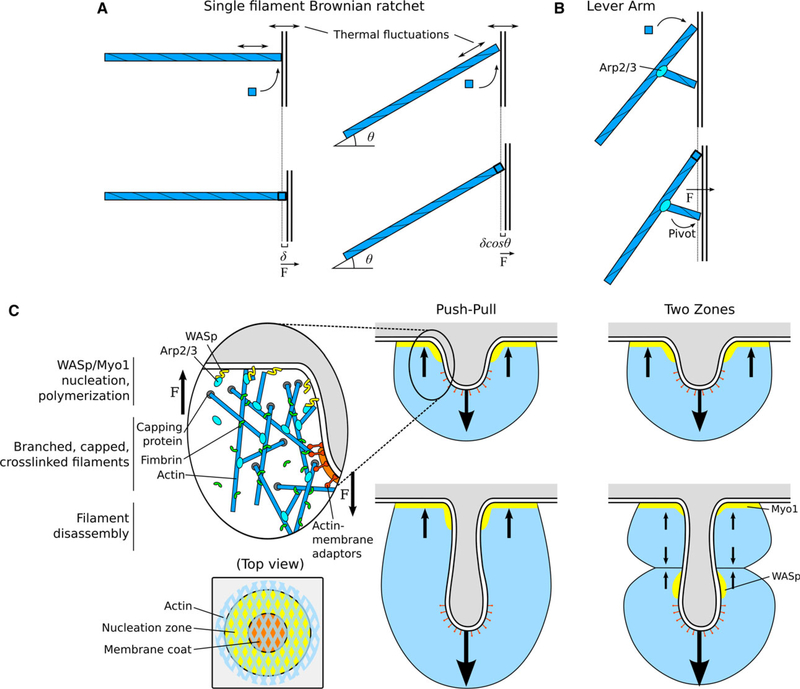
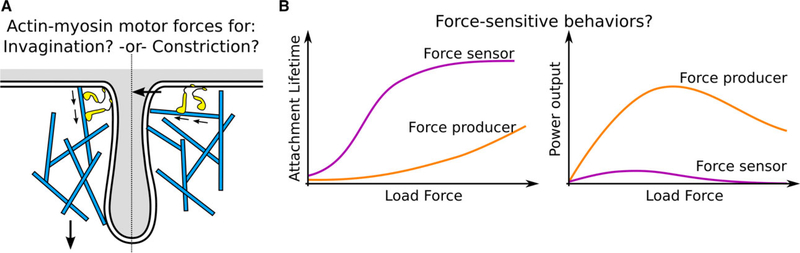
During clathrin-mediated endocytosis (CME), a flat patch of membrane is invaginated and pinched off to release a vesicle into the cytoplasm. In yeast CME, over 60 proteins-including a dynamic actin meshwork-self-assemble to deform the plasma membrane. Several models have been proposed for how actin and other molecules produce the forces necessary to overcome the mechanical barriers of membrane tension and turgor pressure, but the precise mechanisms and a full picture of their interplay are still not clear. In this review, we discuss the evidence for these force production models from a quantitative perspective and propose future directions for experimental and theoretical work that could clarify their various contributions.
Keywords: actin; clathrin; endocytosis; membrane; membrane remodeling; yeast.
© 2018 Federation of European Biochemical Societies.
Figures



References
-
- Gaidarov I, Santini F, Warren RA and Keen JH (1999) Spatial control of coated-pit dynamics in living cells. Nat Cell Biol 1, 1–7. - PubMed
-
- Kaksonen M, Sun Y and Drubin DG (2003) A pathway for association of receptors, adaptors, and actin during endocytic internalization. Cell 115, 475–487. - PubMed
-
- McMahon HT and Boucrot E (2011) Molecular mechanism and physiological functions of clathrinmediated endocytosis. Nat Rev Mol Cell Biol 12, 517–533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
