

A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality
- PMID: 30007418
- PMCID: PMC6086366
- DOI: 10.1016/j.cell.2018.06.035
A Suite of Transgenic Driver and Reporter Mouse Lines with Enhanced Brain-Cell-Type Targeting and Functionality
Abstract
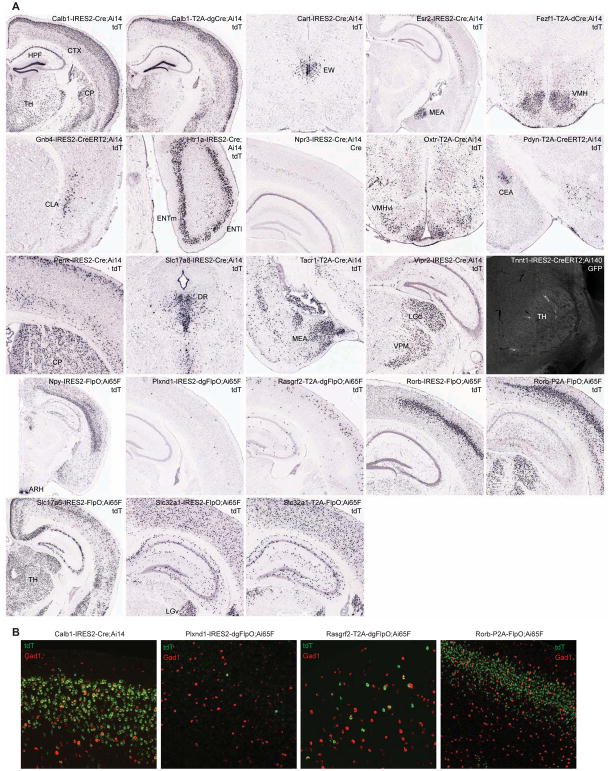
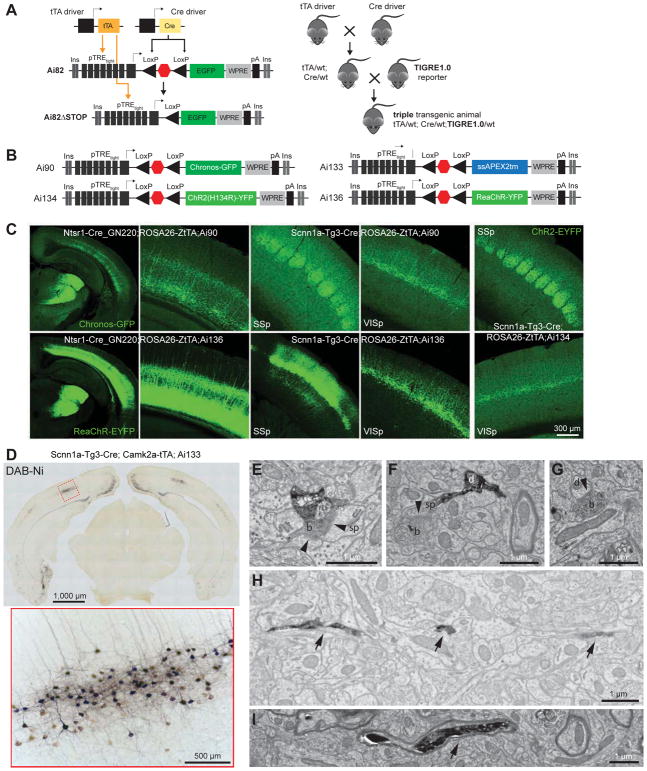
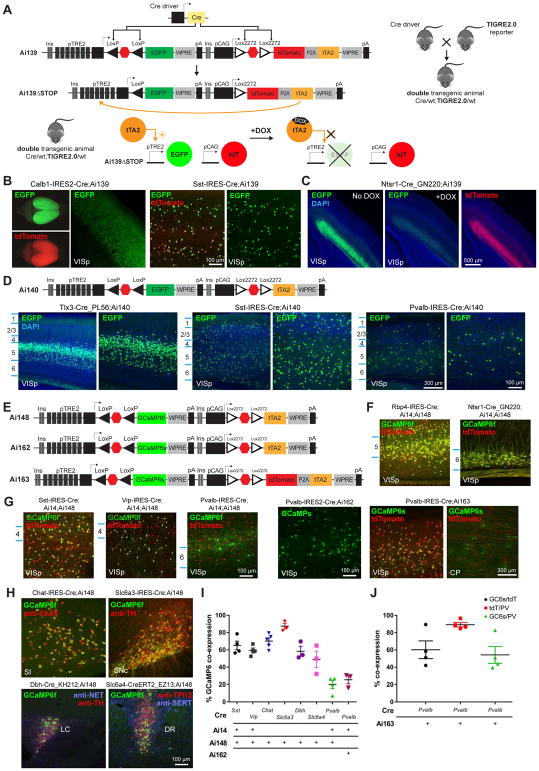
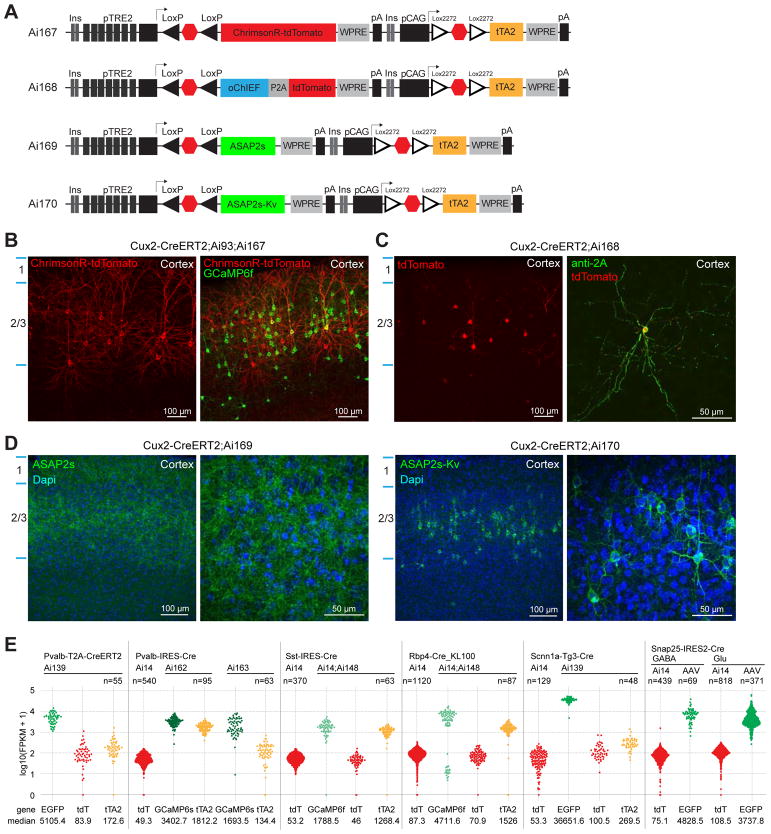
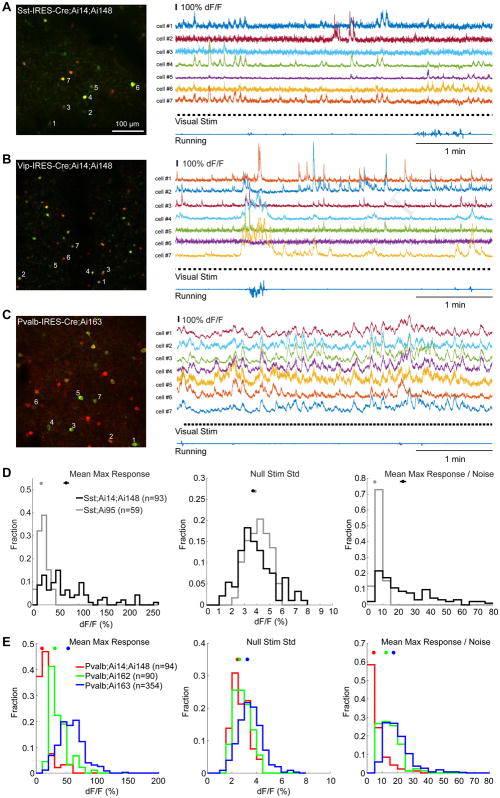
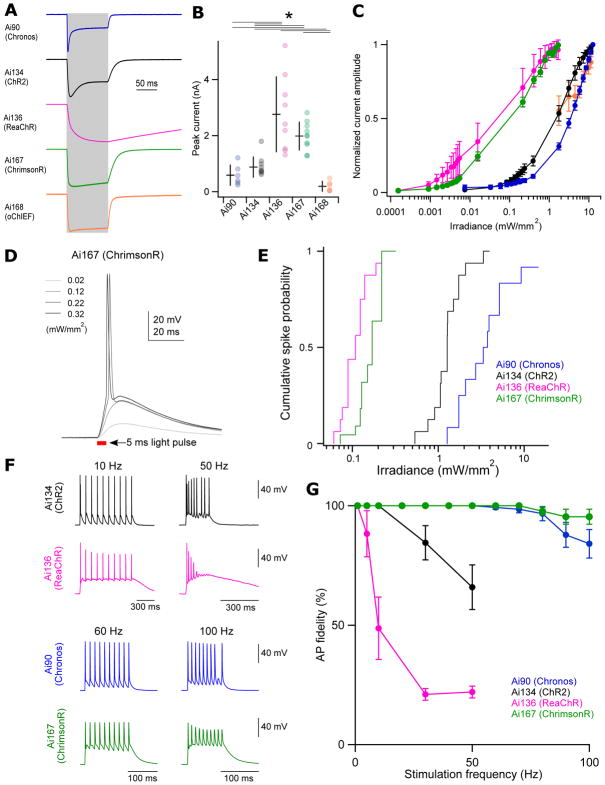
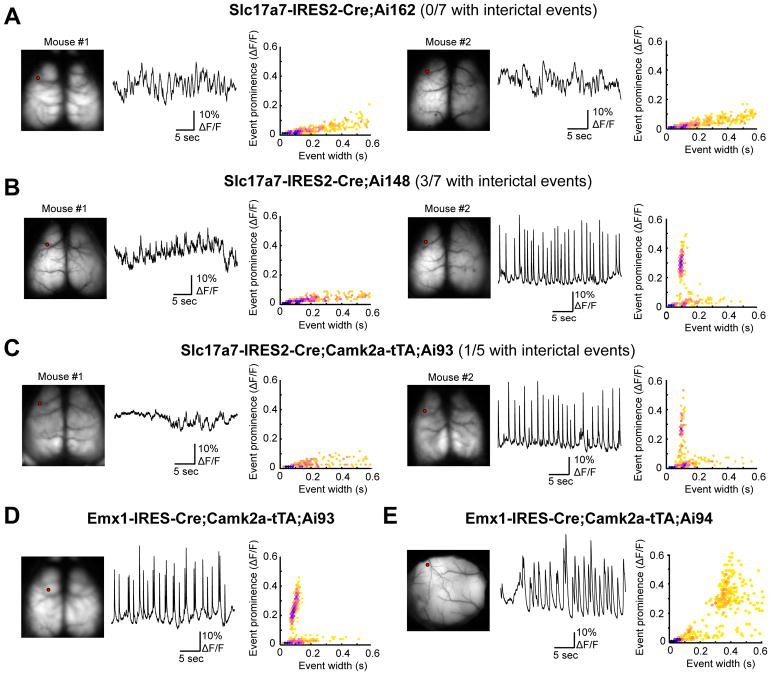
Modern genetic approaches are powerful in providing access to diverse cell types in the brain and facilitating the study of their function. Here, we report a large set of driver and reporter transgenic mouse lines, including 23 new driver lines targeting a variety of cortical and subcortical cell populations and 26 new reporter lines expressing an array of molecular tools. In particular, we describe the TIGRE2.0 transgenic platform and introduce Cre-dependent reporter lines that enable optical physiology, optogenetics, and sparse labeling of genetically defined cell populations. TIGRE2.0 reporters broke the barrier in transgene expression level of single-copy targeted-insertion transgenesis in a wide range of neuronal types, along with additional advantage of a simplified breeding strategy compared to our first-generation TIGRE lines. These novel transgenic lines greatly expand the repertoire of high-precision genetic tools available to effectively identify, monitor, and manipulate distinct cell types in the mouse brain.
Keywords: Cre; Flp; TIGRE; calcium sensor; cell type; channelrhodopsin; optogenetics; reporter; transgenic mice; voltage sensor.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Expansion of the transgenic toolkit in mouse.Nat Methods. 2018 Sep;15(9):651. doi: 10.1038/s41592-018-0124-x. Nat Methods. 2018. PMID: 30171248 No abstract available.
References
-
- Dymecki SM, Ray RS, Kim JC. Mapping cell fate and function using recombinase-based intersectional strategies. Methods in enzymology. 2010;477:183–213. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
