

Marked Diversity of Unique Cortical Enhancers Enables Neuron-Specific Tools by Enhancer-Driven Gene Expression
- PMID: 30008330
- PMCID: PMC7301410
- DOI: 10.1016/j.cub.2018.05.015
Marked Diversity of Unique Cortical Enhancers Enables Neuron-Specific Tools by Enhancer-Driven Gene Expression
Abstract
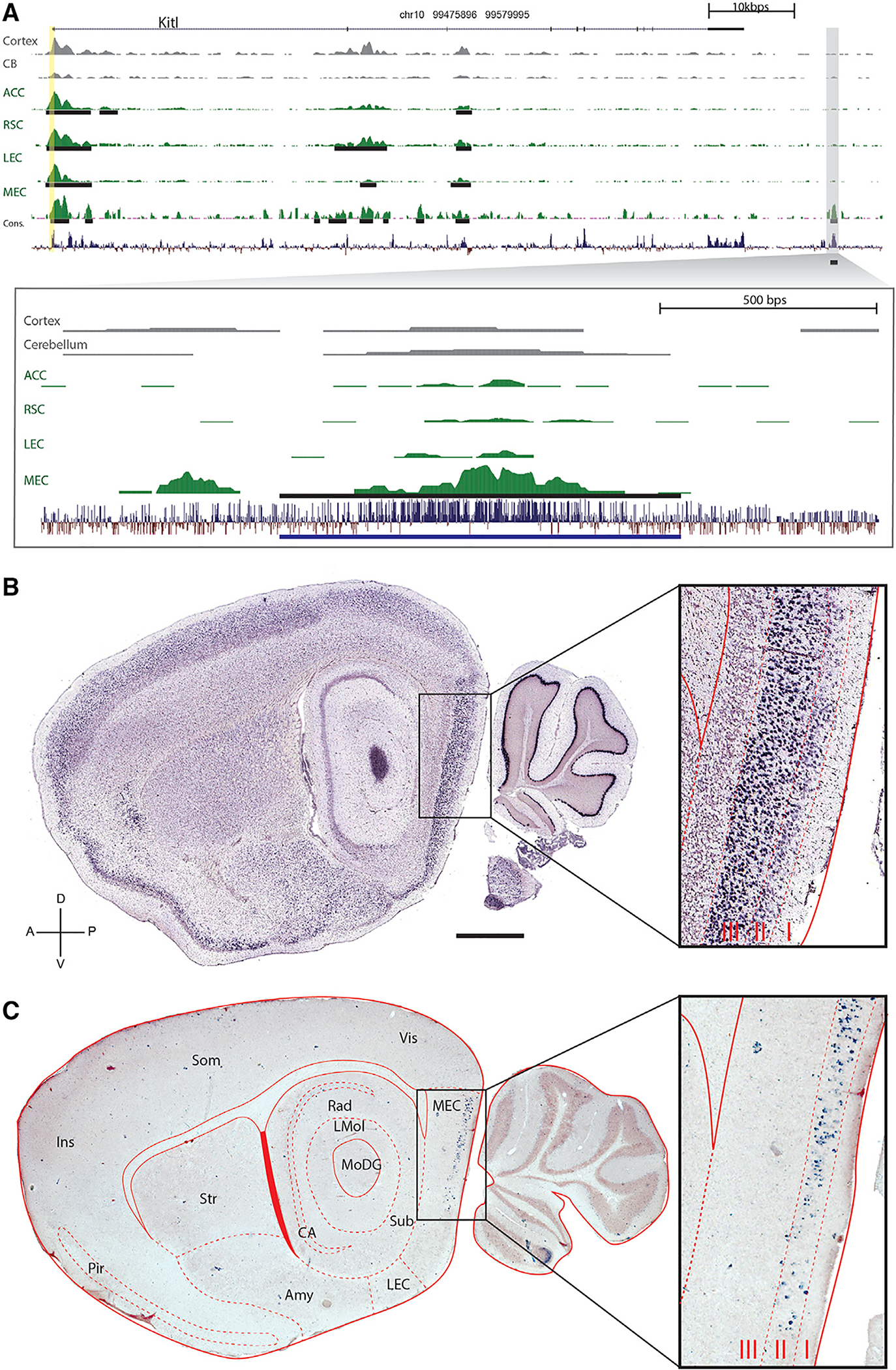
Understanding neural circuit function requires individually addressing their component parts: specific neuronal cell types. However, not only do the precise genetic mechanisms specifying neuronal cell types remain obscure, access to these neuronal cell types by transgenic techniques also remains elusive. Whereas most genes are expressed in the brain, the vast majority are expressed in many different kinds of neurons, suggesting that promoters alone are not sufficiently specific to distinguish cell types. However, there are orders of magnitude more distal genetic cis-regulatory elements controlling transcription (i.e., enhancers), so we screened for enhancer activity in microdissected samples of mouse cortical subregions. This identified thousands of novel putative enhancers, many unique to particular cortical subregions. Pronuclear injection of expression constructs containing such region-specific enhancers resulted in transgenic lines driving expression in distinct sets of cells specifically in the targeted cortical subregions, even though the parent gene's promoter was relatively non-specific. These data showcase the promise of utilizing the genetic mechanisms underlying the specification of diverse neuronal cell types for the development of genetic tools potentially capable of targeting any neuronal circuit of interest, an approach we call enhancer-driven gene expression (EDGE).
Keywords: enhancers; entorhinal cortex; epigenetics; neural circuits; transcriptional control; transgene expression; transgenic animals; transgenic methods; transgenics.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
C.K., J.C., and S.B. are inventors on US Patent Application no. 62/584,282, Appl. Norwegian University of Science and Technology (NTNU), which is related to this work. The authors have no other competing interests to declare.
Figures






References
-
- Zeng H, and Sanes JR (2017). Neuronal cell-type classification: challenges, opportunities and the path forward. Nat. Rev. Neurosci 18, 530–546. - PubMed
-
- Lein ES, Hawrylycz MJ, Ao N, Ayres M, Bensinger A, Bernard A, Boe AF, Boguski MS, Brockway KS, Byrnes EJ, et al. (2007). Genome-wide atlas of gene expression in the adult mouse brain. Nature 445, 168–176. - PubMed
-
- Feng G, Mellor RH, Bernstein M, Keller-Peck C, Nguyen QT, Wallace M, Nerbonne JM, Lichtman JW, and Sanes JR (2000). Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron 28, 41–51. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
