

Morphological Stasis and Proteome Innovation in Cephalochordates
- PMID: 30013013
- PMCID: PMC6071037
- DOI: 10.3390/genes9070353
Morphological Stasis and Proteome Innovation in Cephalochordates
Abstract
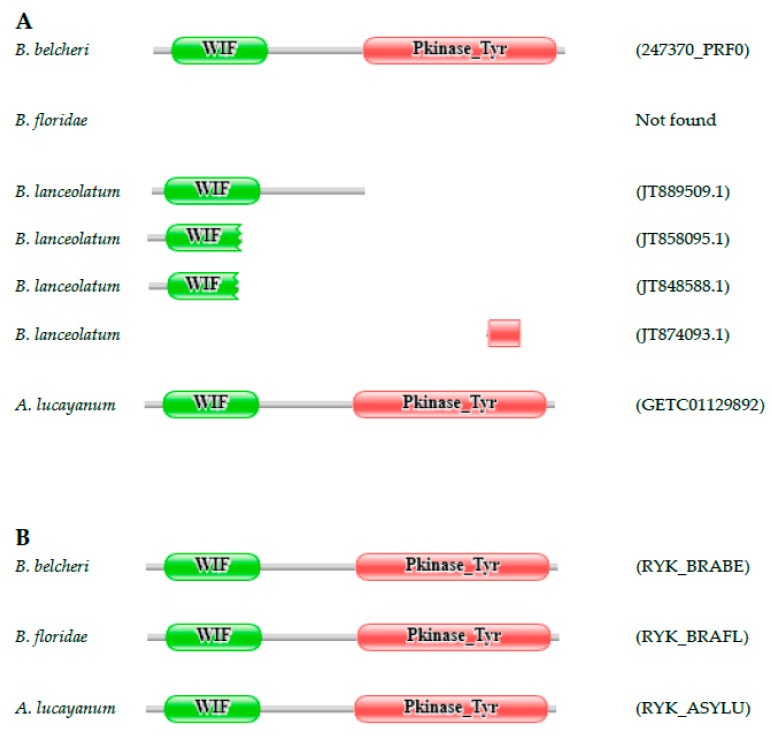
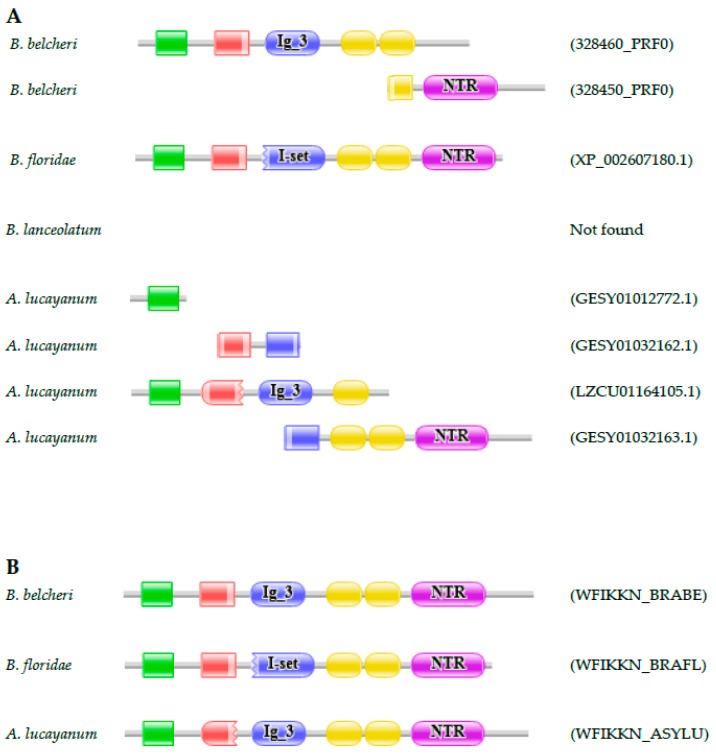
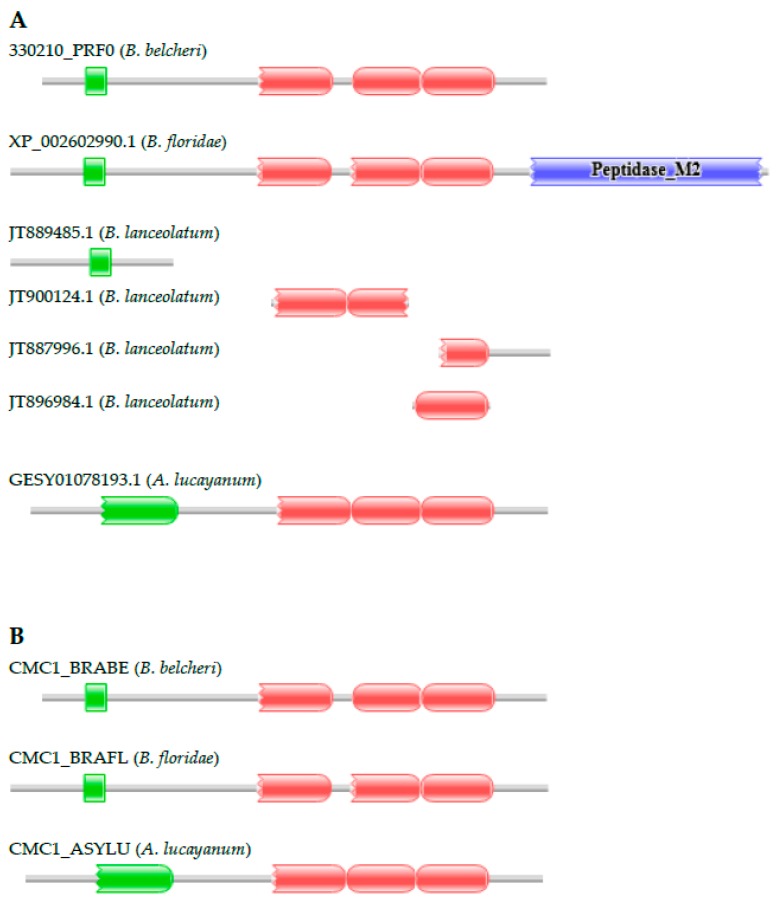
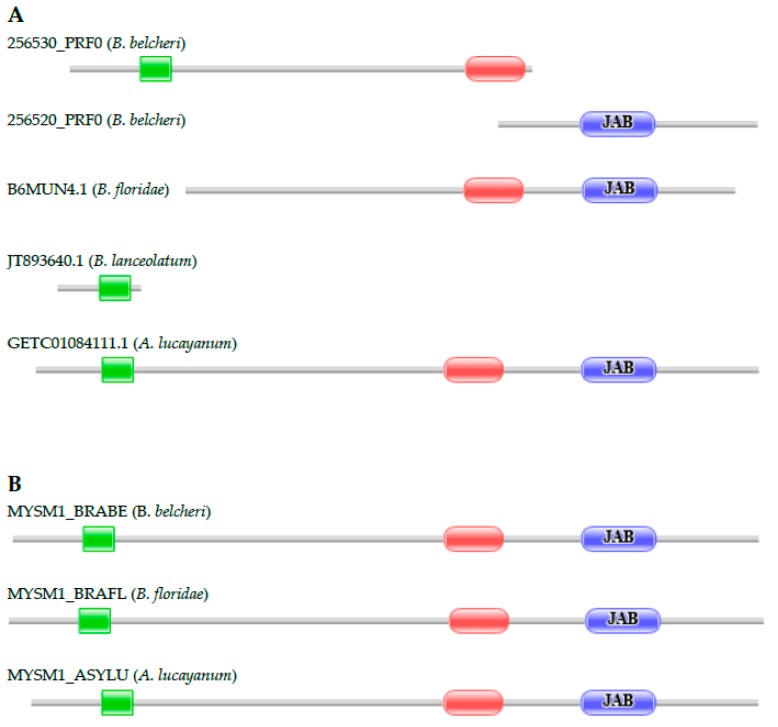
Lancelets, extant representatives of basal chordates, are prototypic examples of evolutionary stasis; they preserved a morphology and body-plan most similar to the fossil chordates from the early Cambrian. Such a low level of morphological evolution is in harmony with a low rate of amino acid substitution; cephalochordate proteins were shown to evolve slower than those of the slowest evolving vertebrate, the elephant shark. Surprisingly, a study comparing the predicted proteomes of Chinese amphioxus, Branchiostoma belcheri and the Florida amphioxus, Branchiostoma floridae has led to the conclusion that the rate of creation of novel domain combinations is orders of magnitude greater in lancelets than in any other Metazoa, a finding that contradicts the notion that high rates of protein innovation are usually associated with major evolutionary innovations. Our earlier studies on a representative sample of proteins have provided evidence suggesting that the differences in the domain architectures of predicted proteins of these two lancelet species reflect annotation errors, rather than true innovations. In the present work, we have extended these studies to include a larger sample of genes and two additional lancelet species, Asymmetron lucayanum and Branchiostoma lanceolatum. These analyses have confirmed that the domain architecture differences of orthologous proteins of the four lancelet species are because of errors of gene prediction, the error rate in the given species being inversely related to the quality of the transcriptome dataset that was used to aid gene prediction.
Keywords: Asymmetron; Branchiostoma; amphioxus; domain architecture; gene prediction; genome annotation; lancelet; proteome; stasis; transcriptome.
Conflict of interest statement
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
Figures

Similar articles
-
An Updated Staging System for Cephalochordate Development: One Table Suits Them All.Front Cell Dev Biol. 2021 May 20;9:668006. doi: 10.3389/fcell.2021.668006. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34095136 Free PMC article.
-
The transcriptome of an amphioxus, Asymmetron lucayanum, from the Bahamas: a window into chordate evolution.Genome Biol Evol. 2014 Sep 19;6(10):2681-96. doi: 10.1093/gbe/evu212. Genome Biol Evol. 2014. PMID: 25240057 Free PMC article.
-
Putative extremely high rate of proteome innovation in lancelets might be explained by high rate of gene prediction errors.Sci Rep. 2016 Aug 1;6:30700. doi: 10.1038/srep30700. Sci Rep. 2016. PMID: 27476717 Free PMC article.
-
Keeping amphioxus in the laboratory: an update on available husbandry methods.Int J Dev Biol. 2017;61(10-11-12):773-783. doi: 10.1387/ijdb.170192ms. Int J Dev Biol. 2017. PMID: 29319123 Review.
-
Gene regulation in amphioxus: An insight from transgenic studies in amphioxus and vertebrates.Mar Genomics. 2015 Dec;24 Pt 2:159-66. doi: 10.1016/j.margen.2015.06.003. Epub 2015 Jun 18. Mar Genomics. 2015. PMID: 26094865 Review.
Cited by
-
Evolution and Structure of Proteins and Proteomes.Genes (Basel). 2018 Nov 28;9(12):583. doi: 10.3390/genes9120583. Genes (Basel). 2018. PMID: 30487453 Free PMC article.
-
Molecular taxonomy confirms that the northeastern Atlantic and Mediterranean Sea harbor a single lancelet, Branchiostoma lanceolatum (Pallas, 1774) (Cephalochordata: Leptocardii: Branchiostomatidae).PLoS One. 2021 May 6;16(5):e0251358. doi: 10.1371/journal.pone.0251358. eCollection 2021. PLoS One. 2021. PMID: 33956890 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
